Материал: Задачи по биохимии
3. Гидролазы. Эти ферменты ускоряют реакции гидролиза органических соединений; обязательным участником этих процессов является вода. В зависимости от характера гидролизуемой связи гидролазы подразделяют на ряд подклассов: эстеразы, гликозидазы, пептидгидролазы и др. Отличительной чертой всех гидролаз является то, что они являются однокомпонентными ферментами.
59
Эстеразы катализируют реакции гидролиза сложноэфирных связей. Приведем примеры:
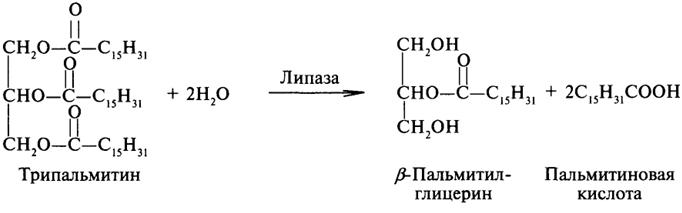
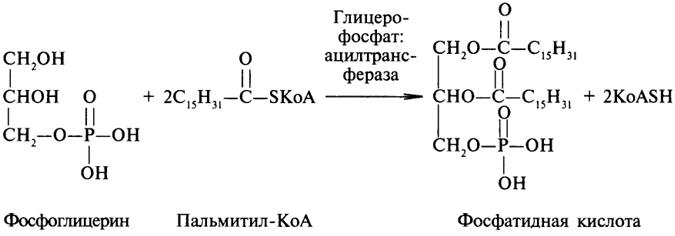
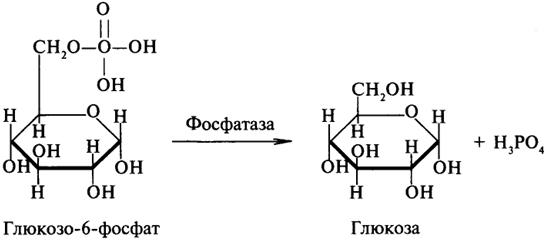
Липаза ускоряет гидролиз внешних сложноэфирных связей в молекуле триглицерида. Особенно широко распространены эстеразы, катализирующие гидролиз сложных эфиров фосфорной кислоты и углеводов. Эти ферменты называются фосфатазами:
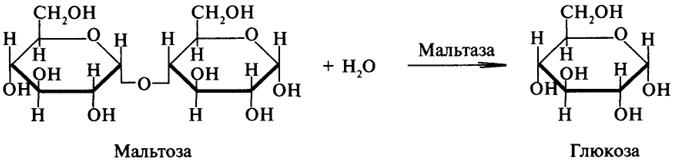
Гликозидазы ускоряют реакции гидролиза гликозидных связей. Примером гликозидазы может служить мальтаза (α-глюкозидаза).
Из гликозидаз, действующих на полисахариды, наиболее распространены амилазы.
Пептид-гидролазы. Ферменты этого подкласса катализируют гидролиз пептидных связей в молекулах пептидов и белков, что можно выразить следующей схемой:
60
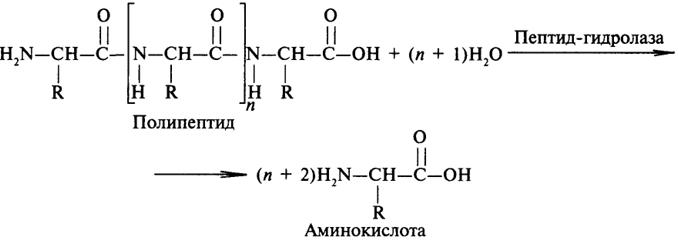
Пептид-гидролазы гидролизуют не все пептидные связи в молекулах белков и пептидов, а только определенные. О специфичности действия пептид-гидролаз речь пойдет в главе "Обмен белков".
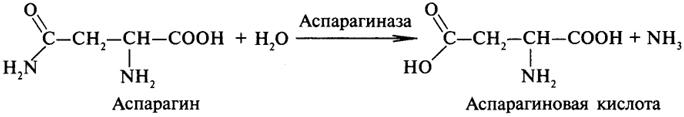
Амидазы ускоряют гидролиз амидов дикарбоновых аминокислот - аспарагина и глутамина.
4. Лиазы. Ферменты этого класса катализируют разнообразные реакции распада и синтеза. В зависимости от того, какая связь расщепляется или, наоборот, образуется, выделяют углерод-углерод, углерод-кислород, углерод-азот лиазы. Приведем примеры процессов, катализируемых ферментами указанных подклассов.
Углерод-углерод лиазы. В природе широко представлены ферменты, ускоряющие декарбоксилирование кето- и аминокислот. Декарбоксилазы или карбокси-лиазы - двухкомпонентные ферменты, коферментом которых является фосфорный эфир витамина B1 - в случае декарбоксилирования кетокислот и витамина В6 - в случае декарбоксилирования аминокислот. Схемы процессов представлены ниже:
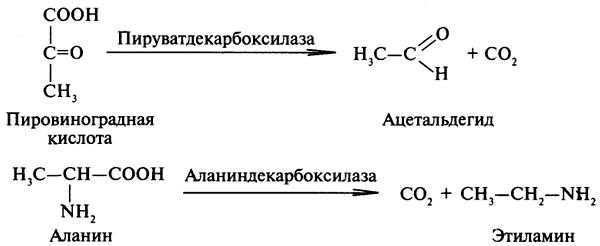
61
Углерод-кислород лиазы (гидролиазы). Ферменты этого подкласса ускоряют реакции гидратации и дегидратации органических соединений.
Эти реакции постоянно идут при распаде и синтезе углеводов и жирных кислот, поэтому гидратазы играют большую роль в жизнедеятельности организмов. Примером может служить фумаратгидратаза, присоединяющая молекулу воды к кратной связи фумаровой кислоты:

Углерод-азот лиазы катализируют реакции прямого дезаминирования некоторых аминокислот; примером может служить аспартат-аммиак-лиаза:

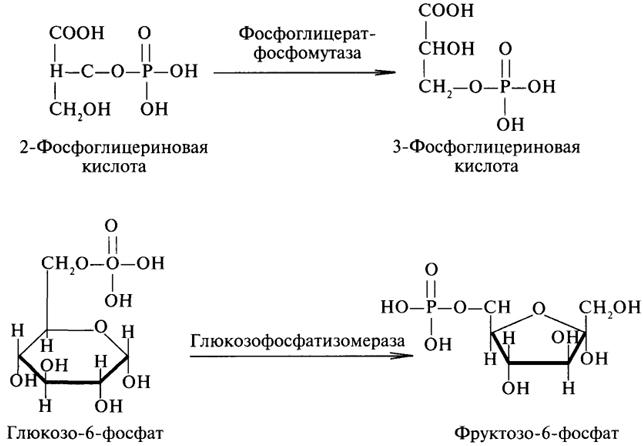
5. Изомеразы. Изомеразы ускоряют процессы превращений одних изомеров органических соединений в другие. Приведем два примера:
6. Лигазы (синтетазы). Ферменты этого класса обеспечивают синтез различных органических соединений. Характерной чертой ферментов этого класса является использование соединений, способных поставлять энергию для осуществления биосинтеза. Одним из таких соединений является аденозинтрифосфорная кислота - АТФ. В качестве примера действия лигазы можно привести синтез щавелевоуксусной кислоты из пировиноградной путем ее карбоксилирования:
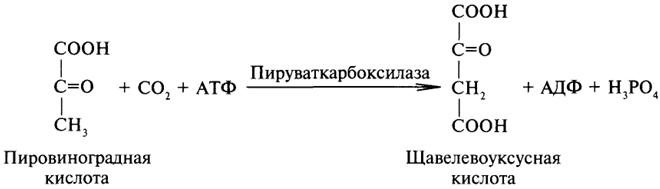
Следует обратить внимание на тот факт, что молекула АТФ не участвует в образовании продуктов реакции, а просто распадается до АДФ и Н3РО4; при этом освобождается энергия, необходимая для осуществления биосинтеза.
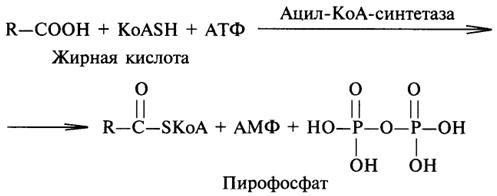
Важной реакцией является образование ацил-коэнзима А (ацил-КоА), которая тоже ускоряется ферментом, относящимся к рассматриваемому классу:
4. Механизм действия ферментов
Ферме́нты, или энзи́мы — обычно белковые молекулы или молекулы РНК (рибозимы) или их комплексы, ускоряющие (катализирующие) химические реакции в живых системах. Реагенты в реакции, катализируемой ферментами, называются субстратами, а получающиеся вещества — продуктами.
-
КФ 1: Оксидоредуктазы, катализирующие перенос электронов, то есть окисление или восстановление. Пример: каталаза, алкогольдегидрогеназа.
-
КФ 2: Трансферазы, катализирующие перенос химических групп с одной молекулы субстрата на другую. Среди трансфераз особо выделяют киназы, переносящие фосфатную группу, как правило, с молекулы АТФ.
-
КФ 3: Гидролазы, катализирующие гидролиз химических связей. Пример: эстеразы, пепсин, трипсин, амилаза, липопротеинлипаза.
-
КФ 4: Лиазы, катализирующие разрыв химических связей без гидролиза с образованием двойной связи в одном из продуктов, а также обратные реакции.
-
КФ 5: Изомеразы, катализирующие структурные или геометрические изменения в молекуле субстрата с образованием изомерных форм.
-
КФ 6: Лигазы, катализирующие образование химических связей C—C, C—S, C—O и C—N между субстратами за счёт реакций конденсации, сопряжённых с гидролизом АТФ. Пример: лигаза
А) понятие энергии активации ферментативных реакций:
Ферменты, действуя как катализаторы, снижают энергию активации. Они повышают общую скорость реакции, не изменяя в сколько-нибудь значительной степени температуру, при которой эта реакция протекает.
Б) активный центр ферментов: определение и структура:
Активный центр — это особая часть молекулы фермента, определяющая её специфичность и каталитическую активность. Активный центр непосредственно осуществляет взаимодействие с молекулой субстрата или с теми её частями, которые непосредственно участвуют в реакции. Характер взаимодействия фермента и субстрата говорит о наличии в активном центре ряда структурных групп, соединяющихся с различными участками субстрата.
В активном центре условно выделяют:
-
каталитический центр — непосредственно химически взаимодействующий с субстратом;
-
связывающий центр (контактная или «якорная» площадка) — обеспечивающий специфическое сродство к субстрату и формирование комплекса фермент-субстрат.
В)
теории образования фермент-субстратного
комплекса по Э. Фишеру и Д. Кошланду: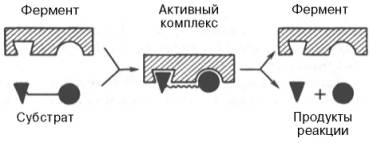
Образование нестойкого фермент-субстратного комплекса согласно теории Э. Фишера «ключ-замок».
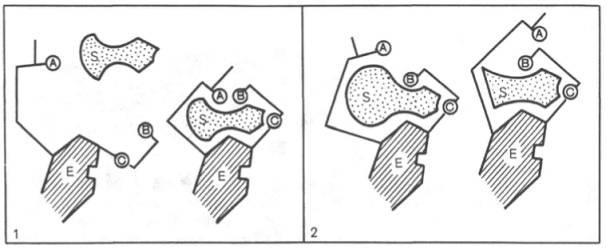
Изменения структуры активного центра фермента, вызванные субстратом, согласно модели «индуцированного соответствия» Д. Кошленда.
Г) уравнение скорости ферментативной реакции, график зависимости скорости ферментативной реакции от концентрации субстрата (кривая субстратного насыщения), константа Михаэлиса и ее биологический смысл:
Уравне́ние Михаэ́лиса — Ме́нтен — основное уравнение ферментативной кинетики, описывает зависимость скорости реакции, катализируемой ферментом, от концентрации субстрата. Уравнение названо в честь физикохимиков Леонора Михаэлиса и Мод Леоноры Ментен. Простейшая кинетическая схема, для которой справедливо уравнение Михаэлиса:
![]()
Уравнение имеет вид:
![]() ,
,
где
-
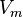 —
максимальная
скорость реакции, равная
—
максимальная
скорость реакции, равная 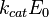 ;
; -
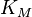 —
константа
Михаэлиса. По определению,
—
константа
Михаэлиса. По определению, 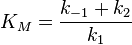 ,
где
,
где 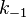 есть
константа скорости реакции распада
фермент-субстратного комплекса на
фермент и исходный субстрат,
есть
константа скорости реакции распада
фермент-субстратного комплекса на
фермент и исходный субстрат, 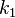 есть
константа скорости реакции образования
фермент-субстратного комплекса и
есть
константа скорости реакции образования
фермент-субстратного комплекса и 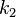 есть
константа скорости реакции распада
фермент-субстратного комплекса на
фермент и продукт (см. ниже вывод
уравнения для скорости реакции).
Константа Михаэлиса численно равна
концентрации субстрата, при которой
скорость реакции составляет половину
от максимальной[1];
есть
константа скорости реакции распада
фермент-субстратного комплекса на
фермент и продукт (см. ниже вывод
уравнения для скорости реакции).
Константа Михаэлиса численно равна
концентрации субстрата, при которой
скорость реакции составляет половину
от максимальной[1]; -
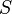 —
концентрация
субстрата.
—
концентрация
субстрата.
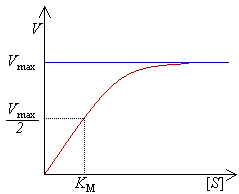
график зависимости скорости ферментативной реакции от концентрации субстрата (кривая субстратного насыщения)
Константа Михаэлиса, Км (Michaelis constant, Km) — кинетический параметр ферментативной реакции, численно равный концентрации субстрата, при которой скорость реакции составляет половину максимальной; К.М. характеризует сродство фермента к субстрату: чем меньше значение К.М., тем сильнее связывание фермента с субстратом.
5. Регуляция действия ферментов
В живой клетке скорость ферментативных реакций находится под строгим контролем, что позволяет каждой метаболической цепочке реакций постоянно изменяться, приспосабливаясь к меняющимся потребностям клетки в продукте.
|
Метаболическая цепь: А, В, С, D - метаболиты, Е1, Е2, Е3, Е4 - ферменты |

В каждой метаболической цепи есть фермент, который задает скорость всей цепочке реакций. Он называется регуляторным ферментом. Существует несколько способов регуляции действия ферментов:
-
изменение активности фермента при его постоянной концентрации;
-
изменение концентрации фермента, обычно в результате ускорения (индукции) или торможения (репрессии) синтеза фермента;
Основные способы регуляции активности ферментов
Аллостерическая регуляция. Фермент изменяет активность с помощью нековалентно связанного с ним эффектора. Связывание происходит в участке, пространственно удаленном от активного (каталитического) центра. Это связывание вызывает конформационные изменения в молекуле белка, приводящие к изменению определенной геометрии каталитического центра. Активность может увеличиться - это активация фермента, или уменьшиться - это ингибирование.
|
Аллостерическая активация фермента |
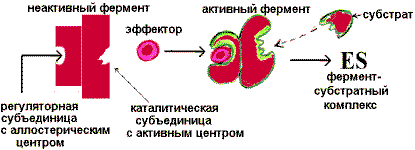
«Сообщение» о присоединении аллостерического активатора передается посредством конформационных изменений каталитической субъединице, которая становится комплементарной субстрату, и фермент «включается». При удалении активатора фермент вновь переходит в неактивную форму и «выключается». Аллостерическая регуляция является основным способом регуляции метаболических путей.
Регуляция активности ферментов путем фосфорилирования-дефосфорилирования . Фермент изменяет активность в результате ковалентной модификации.
|
Регуляция активности липазы |
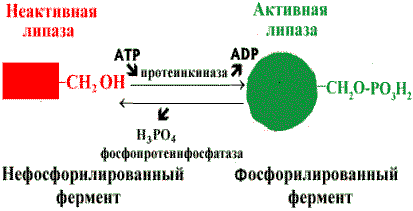
В этом случае фосфатная группа - ОРО32- присоединяется к гидроксильным группам в остатках серина, треонина или тирозина. В зависимости от природы фермента фосфорилирование может его активировать или, наоборот, инактивировать. Реакция присоединения фосфатной группы и ее отщепление катализируют специальные ферменты - протеинкиназы и протеинфосфатазы.
Регуляция путем ассоциации-диссоциации субъединиц в олигомерном ферменте. Этот процесс иногда начинается с ковалентной или нековалентной модификации одной из субъединиц. Например, фермент протеинкиназа в неактивной форме построена как тетрамер R2C2 (R и С - разные субъединицы). Активная протеинкиназа представляет собой субъединицу С, для освобождения которой необходима диссоциация комплекса. Активация фермента происходит при участии cAMP (циклоаденозинмонофосфорная кислота), которая способна присоединиться к субъединице R, после чего изменяется конформация, комплементарность субъединиц R и С и происходит диссоциация комплекса: R2C2 + 2cАМР 2С + 2(R -сАМР) Циклический АМР является продуктом АТР, превращение которой катализирует фермент аденилатциклаза: АТРс АМР + Н4Р2О7
Аденилатциклазная система. Аденилатциклаза и протеинкиназа катализируют взаимосвязанные реакции, которые составляют единую регуляторную систему.
|
Аденилатциклазная система |
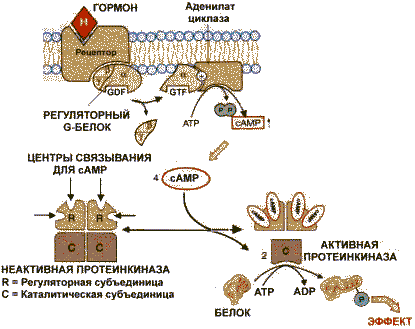
С помощью этой системы в клетку передаются сигналы из внеклеточной среды, и в нужном направлении изменяется метаболизм клетки. Внеклеточным вестником сигнала могут быть разные молекулы, в том числе и гормоны. Эти молекулы не проникают внутрь клетки, но «узнаются» мембранными рецепторами. При активации аденилатциклазы происходят следующие этапы:
|
|
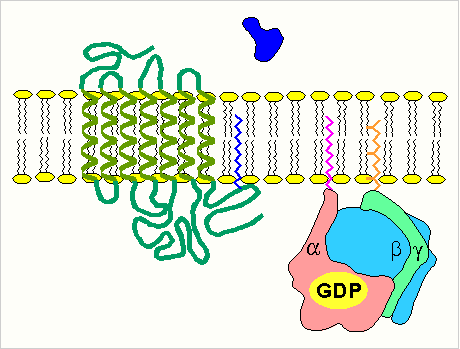
-
изменение конформации рецептора после присоединения к нему сигнальной молекулы и увеличение его сродства к регуляторному G-белку. В результате образуется комплекс рецептора и протомеров G-белка;
-
образование этого комплекса приводит к изменению конформации a -протомера G-белка, который теряет сродство к GDP и происходит замена GDP на GTP. В результате комплекс протомеров G-белка распадается;
-
a -протомер взаимодействует с аденилатциклазой, что ведет к изменению ее конформации и как следствие этого - активации;
-
после этого аденилатциклаза катализирует синтез cAMP, который в свою очередь активирует cAMP-зависимую протеинкиназу. Активация последней связана с диссоциацией комплекса входящих в нее протомеров после присоединения cAMP. Протеинкиназа фосфорилирует соответствующие ферменты, изменяет их активность и, следовательно, скорость метаболизма в клетке.
Активация ферментов путем частичного протеолиза. Некоторые ферменты синтезируются первоначально неактивными и лишь после секреции из клетки переходят в активную форму. Неактивный предшественник называется проферментом. Активация профермента включает модификацию первичной структуры с одновременным изменением конформации. Например, трипсиноген, синтезированный в поджелудочной железе, затем в кишечнике превращается в трипсин путем удаления фрагмента с N-конца: энтеропептидаза трипсиногентрипсин + Val-(Acn) -Lys Расщепление определенных пептидных связей «запускает» новые взаимодействия R-групп по всей молекуле, приводя к новой конформации, в которой R-группы активного центра занимают оптимальное положение для катализа. Нарушения структуры какого-либо фермента, ведущие к снижению его активности, приводят к нарушению метаболических путей, в которых участвует этот фермент. Такие нарушения почти всегда проявляются как болезни. Повреждения ферментов бывают двух типов: наследственные дефекты строения фермента и повреждения, вызванные попадающими в организм токсическими веществами, ингибирующими фермент.