Материал: Вплив антропогенного навантаження на функціонування глутатіонової системи у насінні представників роду Acer L
.4 Функціонування та роль антиоксидантної системи рослин
Різноманітні стресові впливи, яких зазнають материнські
деревні рослини внаслідок дії природних і антропогенних чинників, викликають
зміни фізіолого-біохімічних процесів у насінні [84], зокрема, індукують
відповідні реакції захисних метаболічних систем насіння на вплив полютантів.
Відомо, що ефективний антиоксидантний захист рослин за дії токсикантів різного
походження забезпечує глутатіон-залежна система [19].
.4.1 Структура та властивості глутатіону
Глутатіон являє собою багатофункціональний трипептид,
знайдений в рослинах і тваринах. Це основний небілковий низькомолекулярний
тіол, в більшості організмів (рис. 1.4.1-1.4.2).
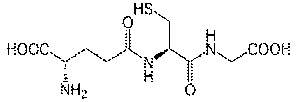
Рис. 1.4.1. Структурна формула глутатіону
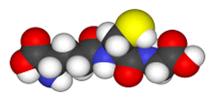
Рис. 1.4.2. Трьохмірна модель молекули глутатіону
Глутатіон бере участь у різноманітних процесах детоксикації, транспорту і метаболізму. Відомо, що він є донором відновлюючих еквівалентів в глутатіон - аскорбонатному циклі (цикл Halliwell Asada). У ході цього процессу відновлена форма глутатіону окислюється для того, щоб відновити дегідроаскарбат (який перетворюється в аскорбат). Перетворення окисненої форми глутатіону в його відновлену форму каталізує глутатіонредуктаза (ГР, КФ 1.8.1.7) [32].
Глутатіон також бере участь у прямій детоксикації пероксиду. У ході цієї реакції відновлений глутатіон (GSH) взаємодіє з пероксидом водню (або іншим органічним пероксидом) з утворенням води, або води та спирту, і димера глутатіону (GSSG).
Ключова роль глутатіону полягає в здатності підтримувати сульфгідрильні группи внутрішньоклітинних білків у правильному стані окислення. Співвідношення TG/GSSGє дуже важливим для гомеостазу клітин і свідчить про здатність рослин протистояти окисному стресу.
Деякі автори [32; 39; 48; 61] припускають, що окислювально-відновний стан глутатіону можна використовувати в якості цінного маркера стресу при вивченні екофізіології рослин.
Глутатіон відповідає за детоксикацію потенційно небезпечних молекул, таких як пестициди, або важкі метали. Кон’югація може відбуватися спонтанно або в присутності GST. Показано велике значення GSTв детоксикації багатьох гербіцидів. Крім того деякі автори припускають, що GST грає важливу роль в процесах фіторимідіації.
Глутатіон також залучається до метаболізму різних сполук, включаючи ароматичні органічні молекули, що відповідають за колір, смак, і аромат, запасні форми відновленої сірки і т.д. Глутатіон не є незамінною речовиною та може синтезуватися з амінокислот L-цистеїну, та L-глутаміновой кислоти і гліцину. Біосинтез відбувається в дві ATP-залежні стадії. На першій стадії синтезується гамма-глутамілцистеїн з L-глутаматом і цистеїном ферментом гаммаглутамілцистеїн синтетазою (або глутаматцистеїн лигазою). Ця реакція є лімітоючою в синтезі глутатіону. На другій стадії фермент глутатіон сінтетаза приєднує залишок гліцину до С-кінцевої групи гама-глутамілцистеїна [22].
Глутатіон бере участь в синтезі лейкотриєнів і є учасником
ферменту глутатіонпероксидази. Як частина глиоксалазної ферментативної системи
глутатіон бере участь у реакції детоксифекації метилглиоксаля, токсичного
побічного продукту метаболізму. Глиоксалаза перетворює метилгліоксаль, і відновлений
глутатіон в лактоілглутатіон. Глеоксалаза гідролізує лактоілглутатіон на
глутатіон і лактат. Глутатіон є субстратом реакції кон’югування і відновлення,
що каталізується глутатіон-s-трансферазою в цитозолі, мікросомах і в
мітохондріях [39].
.4.2 Функції глутатіон-залежних ферментів
Система глутатіону включає в себе три глутатіонзалежних ферменти: глутатіон-пероксидази (GPX), глутатіон-редуктазу (GR), глутатіон-трансферазу (GST).Центральний метаболіт системи - трипептидглутатіон (GSH) глутамілцистеїнілглицин. глутатіон відновлений володіє власною антиоксидантною активністю і виступає в ролі кофактора антиоксидантних ферментів, донора водню, метаболіту і субстрату з ферментами системи, а також з супероксиддисмутазою і каталазою, а також ферментів, що містять тіоловую групу. Глутатіон постійно утворюється в печінці і виділяється в кров, звідки надходить до всіх тканин, крім еритроцитів. Виділяється глутатіон відновлений в жовч. Він бере участь у синтезі білків і нуклеїнових кислот; захищає від активних форм кисню, відновлює та ізомеризує дисульфідні зв'язки, впливає на активність ферментів та інших білків, підтримує функції мембран, виконує деякі коферментні функції, бере участь в обміні ейкозаноїдів; є резервом цистеїну; бере участь у метаболізмі ксенобіотиків; підвищує резистентність клітин до шкідливих впливів; впливає на проліферацію.
Система GSH-глутатіону бере участь в транспорті амінокислот, впливає на функції ниркових мембран і резистентність клітин. Вважають, що система GSH-глутатіону - один із захисних механізмів проти старіння організму [54].
Глутатіон-пероксидаза каталізує реакції, у яких фермент
відновлює пероксид водню до води, а також відновлення органічних
гідропероксидів (RООН) до гидроксипроизводних, і в результаті переходить в
окислену дисульфідну форму GS-SG :
GSН + Н202 = GS-SG + Н20
GSH + RООН = GS-SG + ROН +Н20
Глутатіон-редуктаза - флавопротеїн з простетичною групою
флавинадениндинуклеотидом, складається з двох ідентичних субодиниць.
Глутатіонредуктаза каталізує реакцію відновлення глутатіону з окисленої його
форми GS-SG, а всі інші ферменти глутатіонсинтетаз використовують його:
NADPH + GS-SH = 2NADPH + 2 GSH
Глутатіон-трансфераза (КФ 2.5.1.18) - родина мультифункціональних білків, які використовують GSH для метаболізму гідрофобних речовин. Гомо- або гетеродімери, з молекулярною масою 43-57 кДа, кожна субодиниця має свій незалежний активний центр, який у свою чергу містить два субцентри G і H. Перший взаємодіє з глутатіоном (містить sh-групу, гістидин і аргінін); H субцентр зв'язує гідрофобний субстрат, в ньому міститься гліцин. Глутатіон-трансфераза каталізує реакцію:
+ GSH = HX + GS-SG
Розрізняють глутатіон-трансферази, які взаємодіють з катіонами (у печінці, кишечнику, нирках) і аніонами (головний мозок, селезінка, легені, плацента, еритроцити). Залежно від субстратної специфічності також розрізняють глутатіон-трансферазу, яка взаємодіє з епоксидами, алкенами, алкілами, алканами, арілами, ентеротоксинами.
Максимальна концентрація глутатіон-трансферази знайдена в печінці при оптимумі рН=7,5. В основному глутатіон-трансферази локалізовані у цитозолі і ендоплазматичному ритікулумі, але зустрічається в ядрах та мітохондріях. Ця група ферментів здійснює детоксикацію різних ксенобіотиків або їх мікросомальних метаболітов шляхом трансформації, а також детоксикації пероксидів. Глутатіон-трансферази грають важливу роль в ендогенному метаболізмі: вони зв'язують і транспортують лейкотрієн С4, жовчні кислоти, білірубін, захищають організм від окислювального стресу шляхом відновлення гидропероксидов жирних кислот і нуклеотидів. Глутатіон-трансферази беруть участь в утворенні і метаболізмі гормонів (лейкотрієнів, простагландінів, естрогену) [59].
Глутатіон-трансфераза має 11 ізоформ, здійснює 4 основних типа реакцій;
) Приєднання до субстрату повної молекули глутатіону:
+ GSH = HRSG
) Нукліофільне заміщення:
+ GSH = HX + SG
) Відновлення органічних пероксидів (гідропероксидів жирних
кислот, кумена) до відповідних спиртів:
2GSH + ROOH = ROH + GS-SG + H2O
Цю активність глутатіон-трансферази розглядають як неселенову глутатіон-пероксидаза.
) Ізомеризація (стероїдів, простагландінів). В реакціях
першого та другого типа утворюються тіоефіри (кон'югати), а глутатіон
втрачається. У реакціях третього типа утворюється GS-SG (окислений глутатіон),
який відновлюється глутатіон-редуктазою. У реакціях четвертого типові GSH не
використовується і працюють як кофермент.
.5 Фізіолого-біохімічні особливості насіння
Насіння - основний рослинний репродукційний матеріал як у дикій природі, так і в культурі. Тому інтродукція будь-яких рослин базується переважно на насіннєвому розмноженні, за винятком тих, що не дають насіння чи здатні тільки до вегетативного розмноження. У процесі еволюції насіння набуло ряду властивостей, які дають йому можливість зберігати життєздатність до того часу, доки не настануть сприятливі для проростання і утворення нових рослин умови.
Однією з таких особливостей є здатність різко змінювати інтенсивність метаболізму від процесів формування зародка та інших частин насінини і відкладення запасних речовин до гідролізу цих речовин і використання їх на ріст зародка при проростанні. Дослідження проростання насіння певною мірою забезпечує успіх рослинництва, в тому числі інтродукції, але саме труднощі з насінництвом часто затримують процес введення нових корисних рослин в культуру [70].
Насіння рослин характеризується цілим рядом морфологічних, біохімічних і фізіологічних особливостей, серед яких вирізняється його фізіологічна неординарність, тобто різноманіття вимог до умов проростання, різний термін настання його фізіологічної зрілості.
Широкий діапазон умов, що необхідні для проростання насіння, сприяє ефективному використанню території, виживанню та співіснуванню різних видів рослин. Для дикорослих видів фізіологічна різноякісність насіння є цінним пристосуванням, за допомогою якого реалізується генетичний потенціал виду, його збереження і поширення [31].
Особливості проростання насіння дикорослих видів рослин на даний час вивчені недостатньо. Для насіння більшості рослин, ареали яких розташовані в помірних широтах, властивий більш або менш тривалий період спокою, завдяки якому утворюються грунтові насіннєві банки, здатні забезпечити резерв для відтворення потомства протягом кількох років. Спокій насіння є однією з складових різноякісності насіння і зумовлений рядом причин, серед яких - фізіолого-біохімічний стан і рівень морфологічного розвитку зародка [46; 55].
До фізіологічних (біологічних) властивостей насіннєвих мас відносять інтенсивність дихання, здатність до проростання та енергію проростання насіння. Ці властивості визначають стан насіннєвої маси під час зберігання і напрямок використання насіння.
Інтенсивність дихання суттєво впливає на процес зберігання насіння та його якість. Якість насіння буде тим вищою, чим нижча його інтенсивність дихання, яка в першу чергу залежить від вологості насіння. Чим вища вологість, тим вища інтенсивність дихання і нижчий її температурний оптимум.
Інтенсивність дихання насіннєвих мас залежить не лише від дихання самого насіння, а від дихання мікроорганізмів, що заселяють поверхню насіння та домішок. Висока інтенсивність дихання супроводжується виділенням тепла, збільшенням температури насіння та активності ферментів, що спричиняє високу швидкість окиснювальних процесів і в кінці кінців може викликати самозігрівання насіння.
Найбільш оптимальним методом запобігання високій інтенсивності дихання є зберігання насіння в сухому стані.
Проростання насіння відбувається за сприятливих умов.
Проростанню насіння передує набрякання - збільшення розмірів внаслідок
поглинання певної кількості води.Швидкість набрякання та кількість поглинутої
води залежать від віку насіння: чим воно молодше, тим швидше набрякає й більше
поглинає води [53].
.5.1 Ортодоксальне насіння
Процес зневоднення розглядається як необхідна подія життєвого
циклу у видів, які продукують так зване ортодоксальне насіння. Під час розвитку
ортодоксального насіння на материнській рослині в ньому відбувається втрата
води, що збігається з набуванням зародками стійкості до зневоднення. Зазвичай
втрачається понад 80% води, в результаті активність метаболічних процесів різко
знижується, а насіння переходить у стан спокою [20; 61]. Дуже важливий той
факт, що разом зі стійкістю до зневоднення насіння набуває неспецифічну
стійкість і до інших стрессорів навколишнього середовища.
.5.2 Рекальцітрантне насіння
Поняття «рекальцітрантність» (в перекладі «непослушность») стало використовуватися з 1980 р. відносно зрілого насіння, яке гине при висиханні, на відміну від звичайного, стійкого до висихання ортодоксального насіння. Вже в період дозрівання обидва типи насіння помітно різняться за водним статусом. Під час дозрівання обидва види насіння поступово втрачають воду, при цьому ортодоксальне насіння в кінці періоду дозрівання висихає до вологості близько 10%.У такому стані воно може зберігатися багато років, не втрачаючи життєздатності. Така властивість ортодоксального насіння отримало назву «стійкість до висихання». На відміну від нього, втрата води рекальцітрантним насінням призводить до втрати схожості і загибелі - тому його вважають чутливим до висихання.
Вивчення фізіології рекальцитрантного насіння досі було зосереджено на природі його чутливості до висихання[27; 74], тобто на його принципову відмінність від ортодоксального насіння. Встановлено, що причиною загибелі при висиханні є недостатня активність антиоксидантного захисту [89], що призводить до накопичення активних форм кисню в токсичних концентраціях. Однак практично ніким не вивчалася сама фізіологія рекальцитрантності, пов'язана, у першу чергу, з підтриманням високої оводненості і життєздатності.
Рекальцітрантність насіння найбільше поширена в районах з
помірно-вологим тропічним кліматом, особливо в так званих дощових лісах. Там
зріле насіння відразу падає у вологий грунт і не відчуває нестачі вологи, що
дозволяє йому тут же прорости. З цієї причини насінини не мають стійкості до
висихання. Їх рекальцітрантність дозволяє їм успішно скоротити період часу між
дозріванням і проростанням, що сприяє швидкому відновленню виду.
Рекальцітрантне насіння характерне також і для рослин субтропіків, а також для
місць зростання з сезонними змінами клімату, з високою вологістю в період
дозрівання і проростання насіння [92].
2. Матеріал і методи досліджень
.1 Об’єкти досліджень
Досліджувані нами рослини належать до відділу Квіткові, або Покритонасінні (Magnoliophyta, або Angiospermae ), класу Магноліопсиди, або Дводольні (Magnoliopsida, або Dicotyledones), родини Кленові (Aceraceae), роду Клен (Acer L.).
Клен несправжньоплатановий, або явір (A. pseudoplatanus L.),-
струнке дерево до 40 м заввишки, діаметром до 12 м, з густою, гарною,
шатровидною кроною у поодиноких екземплярів, широкоциліндричною - у
насадженнях. Стовбур з попелясто-сірою корою, яка розтріскується і
відшаровується великими шматками у старих екземплярів, подібно до платану
оголюючи молоду кору світло-сірого або рожевого кольору, внаслідок чого стовбур
стає дуже декоративним. Пагони голі, світлі, буро-жовті.

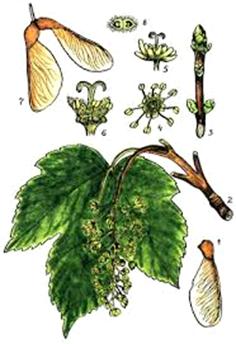
Рис. 2.1.1. Acer pseudoplatanus L.
Листки 3-5-лопатеві, до 17 см, зверху темно-зелені, тьмяні, голі, знизу сизуваті або білуваті, іноді пурпурово-червоні, з неглибокими, загостреними, крупнозубчастими лопатями. Суцвіття - вузькі, багатоквіткові кисті до 16 см завдовжки. Тривалість цвітіння 10-15 днів.
Росте швидко, але повільніше клена гостролистого. Погано мириться з сухими і надмірно вологими грунтами, не переносить засолення, досить тіньовитривалий і теплолюбний.
Форми ефектніше виглядають на освітлених ділянках. Завдяки потужному зростанню, ефектною овальної кроні і жовто-зеленому листі красивої форми, довго зберігає своє забарвлення, належить до групи високодекоративних порід для групових, рядових і одиночних посадок. У культурі дуже давно.
Насіння зберігають у щільно закупорених судинах або в пластикових запаяних пакетах в неопалюваному приміщенні. При цьому схожість зберігається 2 роки. Лабораторна схожість насіння 85 %, ґрунтова - 75%. Насіння висівають відразу ж після збору (сходи з'являються навесні наступного року) або стратифікують спочатку в торфі 45 днів при - 3°С, а потім під снігом протягом 45 днів. Глибина загортання 4-5 див. Клен псевдоплатановий чудовий своєю стійкістю до вітру, міським забруднень і солі, що робить його популярним для вирощування в містах, уздовж доріг, посипати сіллю взимку, і на морському узбережжі.