Материал: В-Л-Быков-Цитогогия_и_общая_гистология
Вместе с тем, в действительности, активные процессы эндоцитоза и экзоцитоза не приводят к существенным изменениям площади поверхности плазмолеммы, так как они уравновешиваются формированием экзоцитозных и эндоцитозных пузырьков, соответственно, компенсирующим происходящую потерю мембраны или ее увеличение за счет противоположно направленного процесса. Эти явления отражают постоянно происходящий в клетке круговорот мембран, который получил название "мембранного конвейера".
Мембранные рецепторы являются преимущественно гликопротеинами, которые расположены на поверхности плазмолеммы клеток и обладают способностью высокоспецифически связываться со своими лигандами. Они выполняют ряд функций:
(1)регулируют проницаемость плазмолеммы, изменяя конформацию белков и ионных каналов;
(2)регулируют поступление некоторых молекул в клетку;
(3)действуют как датчики, превращая внеклеточные сигналы во внутриклеточные;
(4)связывают молекулы внеклеточного матрикса с цитоскелетом, эти рецепторы, называемые интегринами, играют важную роль в формировании контактов между клетками и клеткой и компонентами межклеточного вещества.
Рецепторы, связанные с каналами, взаимодействуют с сигнальной молекулой (нейромедиатора), которая временно открывает или закрывает воротный механизм, в результате чего инициируется или блокируется транспорт ионов через канал.
Каталитические рецепторы включают внеклеточную часть (собственно рецептор) и цитоплазматическую часть, которая функционирует как протеинкиназа (посредством таких рецепторов на клетки воз-действуют инсулин и некоторые факторы роста).
Рецепторы, связанные с G-белками - трансмембранные белки, ас-
социированные с ионным каналом или ферментом, - состоят из рецептора, взаимодействующего с сигнальной молекулой (первый посредник), и G-белка (гуанозин трифосфат-связывающего регуляторного белка), включающего несколько компонентов), который передает сигнал на связанный с мембраной фермент (аденилат циклазу) или ионный канал, вследствие чего активируется второй внутриклеточный посредник - чаще всего циклический АМФ (цАМФ) или Са2+. Около 80% всех гормонов и нейромедиаторов действуют через рецепторы, связанные с эффекторными механизмами посредством G-белков.
В составе плазмолеммы находятся интегрины, называемые клеточными адгезионными молекулами (KAM) - трансмембранные белки, слу-
- 41 -
жащие рецепторами для внеклеточных фибриллярных макромолекул фибронектина и ламинина (см. рис. 10-9). Фибронектин связывается с клетками
имолекулами внеклеточного матрикса (коллагеном, гепарином, фибрином). Таким образом, фибронектин играет роль адгезионного мостика между клеткой
икомпонентами межклеточного вещества. Между тем, внутриклеточная часть молекулы интегрина через ряд друих белков (талин, винкулин и α-актинин) связана с цитоскелетом.
Поверхностный аппарат клетки выделяется некоторыми авторами. которые рассматривают его как структурно и функционально единое образование, состоящее из трех компонентов: (1) надмембранного комплекса (гликокаликса), (2) плазмолеммы и (3) подмембранного комплекса (см. рис. 3- 17). Первые два компонента описаны выше. Подмембранный комплекс образован специализированной периферической частью цитоплазмы, прилежащей к плазмолемме (кортикальный слой) и содержащей элементы цитоскелета (см. ниже), преимущественно актиновые микрофиламенты. Более глубоко располагаются промежуточные филаменты и микротрубочки. Благодаря сокращению сети микрофиламентов, связанных с белками плазмолеммы, происходят изменения формы клетки и ее отдельных участков, формирование псевдоподий, выростов, перемещение клетки в пространстве.
СИНТЕТИЧЕСКИЙ АППАРАТ КЛЕТКИ
Синтетический аппарат клетки включает органелльг, участвующие в синтезе различных веществ, которые могут в дальнейшем использоваться самой клеткой или выделяться ею во внеклеточное пространство. Деятельность синтетического аппарата клетки, располагающегося в ее цитоплазме, контролируется ядром благодаря активности находящихся в нем генов. В синтетический аппарат входят рибосомы, эндоплазматическая сеть (ЭПС) и комплекс Гольджи.
Рибосомы
Рибосомы - мелкие (диаметр - 15-30 нм) плотные немембранньге органеллы, обеспечивающие синтез белка путем соединения аминокислот в попипептидные цепочки. Информация о синтезе приносится к рибосомам информационной РНК (иРНК), которая образуется в ядре в ходе считывания (транскрипции) фрагментов генетической информации с ДНК. Синтетически активная клетка содержит несколько миллионов рибосом (например, в клетке печени их число составляет 107), на которые приходится около 5% ее сухой массы.
- 42 -
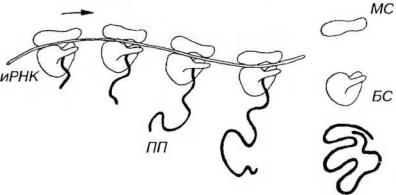
Каждая рибосома состоит из двух асимметричных субъединиц: малой, связывающей РНК, и большой, катализирующей образование пептидных цепей (рис. 3-6). По форме малая субъединица напоминает телефонную трубку, большая - ковш. Субъединицы образованы рибосомальными РНК (рРНК), на которые приходится около 50% их массы, и особыми белками (до 80 различных видов). Первые образуются в ядрышке, белки же синтезируются в цитоплазме, после чего транспортируются в ядро, где связываются с рРНК. В дальнейшем субъединицы по-отдельности через ядерные поры направляются из ядра в цитоплазму, где они участвуют в синтезе белка.
Рис. 3-6. Синтез белка на полирибосоме. Молекула синтезируемого полипептида (ПП) удлиняется по мере движения рибосом (Р), образующих полирибосому, по иРНК (направление показано стрелкой). По завершении синтеза ПП отделяется от Р, которые диссоциирует на две субъединицы - малую (МС) и большую (БС).
Рибосомы могут встречаться в цитоплазме поодиночке (в этом случае они функционально неактивны) или формировать скопления, которые называются полирибосомами (полисомами). В последних отдельные рибосомы (в количестве 3-30) удерживаются общей нитью иРНК толщиной 1.5 нм (см. рис. 3-6). Информация, переносимая иРНК, кодирует последовательность аминокислот в белке соответствующей последовательностью нуклеотидов. Рибосомы переводят (транслируют) эту генетическую информацию в реальную последовательность аминокислот в ходе белкового синтеза.
Функционально неактивные (нетранслирующие) рибосомы постоянно обмениваются своими субъединицами; их сборка происходит в начале синтеза белка, а по завершении синтеза одного полипептида они вновь обратимо диссоциируют.
- 43 -
Синтез белка рибосомой (см. рис. 3-6) начинается со связывания малой субъединицы с участком иРНК; далее рибосома передвигается вдоль цепи иРНК, причем на каждом этапе происходит специфическое присоединение к рибосоме молекулы транспортной РНК (тРНК), антикодон которой комплементарен соответствующему кодону иРНК. В полипептид включается около 20 аминокислот в 1 секунду; белковая молекула среднего размера синтезируется за 20-60 с. Когда образование белковой цепочки завершается, субъединицы диссоциируют, освобождаясь от иРНК. Пока продолжается синтез белка данной рибосомой, новая рибосома занимает освобождающееся на иРНК место. По этой причине активно транслируемая иРНК находится в полисомах. Средняя продолжительность существования синтезированной белковой молекулы варьирует от нескольких минут до нескольких месяцев и даже лет, составляя в среднем около 2 сут.
Белки, которые после синтеза остаются в гиалоплазме (цитоплазматическом матриксе) клетки и далее используются ею, обычно синтезируются на свободных полисомах. Полисомы, которые своими большими субъединицами прикреплены к мембранам ЭПС, синтезируют белки, накапливающиеся в просвете цистерн ЭПС и в дальнейшем либо секретируемые клеткой, либо запасаемые ею внутри гранул (например, лизосомальные ферменты). На полисомах, связанных с мембранами ЭПС, синтезируется также большая часть интегральных мембранных белков. Будет ли белок синтезироваться на ЭПС или на свободных полисомах, зависит от характера начально образуемого отдела полипептидной цепи (сигнальной последовательности или пептида).
Присутствие значительного числа рибосом в цитоплазме клеток, активно синтезирующих белок, придает ей при исследовании на светооптическом уровне базофилию.
Эндоплазматическая сеть
Эндоплазматическая сеть (ЭПС) - органелла, обеспечивающая синтез углеводов, липидов и белков, а также начальные посттрансляционные изменения последних. Она имеет мембранное строение и состоит из системы уплощенных, удлиненных, трубчатых и везикулярных образований. Название органеллы обусловлено характером связи этих элементов друг с другом, образующих в цитоплазме непрерывную трехмерную сеть, элементы которой лишь на отдельных срезах могут иметь вид изолированных структур. Мембрана ЭПС тоньше, чем плазмолемма и содержит более высокую концентрацию белка, что связано с наличи-
- 44 -
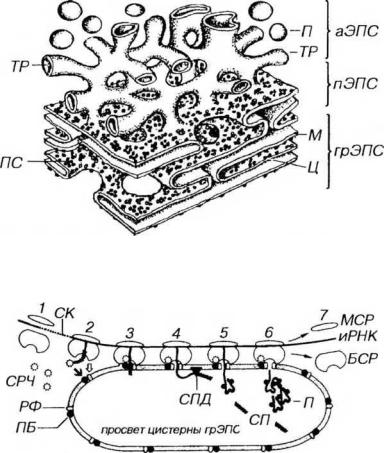
ем в ней многочисленных ферментных систем. Степень развития ЭПС и особенности ее строения варьируют в различных клетках и зависят от их функции. Выделяют две разновидности ЭПС: гранулярную ЭПС (грЭПС) и гладкую, или агранулярную ЭПС (аЭПС), которые связаны друг с другом в области перехода, называемой переходной (транзиторной) ЭПС (рис. 3-7).
Рис. 3-7. Эндоплазматическая сеть. грЭПС: ПС - полисомы, М - мембрана, Ц - цистерны; аЭПС: ТР - трубочка, П - пузырьки; пЭПС - переходная ЭПС.
Рис. 3-8. Синтез белка на гранулярной эндоплазматической сети. БСР - большая субъединица рибосомы, МСР - малая субъединица рибосомы, РФ - рибофорины, СРЧ - сигнал-распознающая частица, ПБ - причальный белок, СК - сигнальные кодоны (иРНК), СП - сигнальный пептид, СПД - сигнальная пептидаза, П - пептид (продукт синтеза). Светлая стрелка - связывание БСР с РФ, темная стрелка - связывание СРЧ с ПБ.
- 45 -