Материал: В-Л-Быков-Цитогогия_и_общая_гистология
Гранулярная ЭПС обеспечивает (1) биосинтез всех мембранных белков и белков, предназначенных для экспорта из клетки, и (2) начальное гликозилирование и посттрансляционные изменения белковых молекул. Гранулярная ЭПС образована уплощенными мембранными цистернами и трубочками, на наружной поверхности которых располагаются рибосомы и полисомы, придающие мембранам зернистый (гранулярный) вид (см. рис. 3-7 и 3-8), что и отражено в названии органеллы. Мембраны грЭПС содержат особые белки, которые обеспечивают (1) связывание рибосом и (2) уплощение цистерн. Полость грЭПС содержит рыхлый материал умеренной плотности (продукты синтеза) и сообщается с перинуклеарным пространством (см. ниже). Благодаря грЭПС происходит отделение (сегрегация) вновь синтезированных белковых молекул от шалоплазмы.
Синтез белка на грЭПС начинается на свободных полисомах, которые в дальнейшем связываются с мембранами ЭПС (см. рис. 3-8). На первом этапе взаимодейстия иРНК с рибосомами происходит образование особого сигнального пептида (длиной 20-25 аминокислот), связывающегося с рибонуклеопротеидным комплексом - сигнал-распознающею частицей (СРЧ). Присоединение СРЧ к сигнальному пептиду угнетает дальнейший синтез белка до тех пор, пока комплекс СРЧ-полисома не свяжется со специфическим рецептором на мембране ЭПС - причальным белком (docking protein в англоязычной литературе). После связывания с рецептором СРЧ отделяется от полисом, что разблокирует синтез белковой молекулы. В мембране грЭПС имеются интегральные рецепторные белки рибофорины, обеспечивающие прикрепление больших субъединиц рибосом. Эти белки не диффундируют в область аЭПС и формируют гидрофобные каналы в мембране, служащие для проникновения вновь синтезированной белковой цепочки в просвет грЭПС, что, наряду с рибофоринами, способствует удержанию рибосом на поверхности мембран грЭПС.
В просвете грЭПС сигнальный пептид отщепляется особым ферментом сигнальной пептидазой, которая располагается на внутренней поверхности мембраны. В ходе продолжающейся трансляции внутри цистерны грЭПС накапливается белок, который приобретает вторичную и третичную структуру, а также подвергается начальным посттрансляционным изменениям - гидроксилированию, сульфатированию и фосфорилированию. Наиболее важным из этих изменений является гликозилирование - присоединение к белкам олигосахаридов с образованием гликопротеинов, которое происходит перед секрецией или транспортом большинства белков к другим участкам внутри клетки (комплексу Гольджи, лизосомам или плазмолемме). В отличие от них, растворимые бел-
- 46 -
ки гаалоплазмы не гликозилированы. Гликозилирование обеспечивается связанным с мембраной ферментом гликозилтрансферазой, переносящим олигосахарид.
Хотя грЭПС присутствует во всех клетках (за исключением спермиев), степень ее развития существенно варьирует. Она особенно хорошо развита в клетках, специализирующихся на белковом синтезе, например, в эпителиальных железистых клетках ацинусов поджелудочной железы (вырабатывающих пищеварительные ферменты), фибробластах (синтезирующих коллаген и ряд других белков), плазматических клетках (продуцирующих иммуноглобулины). Для вех этих клеток характерна выраженная базофилия цитоплазмы в области расположения элементов грЭПС. В нейронах отдельным компактным скоплениям цистерн грЭПС на светооптическом уровне соответствуют очерченные участки базофилии цитоплазмы, которые в совокупности называются хроматофильной субстанцией или тельцами Ниссля.
Агранулярная (гладкая) ЭПС представляет собой трехмерную замкнутую сеть мембранных анастомозирующих трубочек, канальцев, цистерн и пузырьков диаметром 20-100 нм, на поверхности которых рибосомы отсутствуют (см. рис. 3-7), что определило ее название. Соответственно, на мембранах аЭПС отсутствуют рецепторы, связывающие субъединицы рибосом (рибофорины). Предполагают, что аЭПС образуется в результате формирования выростов грЭПС, мембрана которых утрачивает рибосомы.
Функции аЭПС включают: (I) синтез липидов, в том числе мембранных (ферменты липидною синтеза располагаются на наружной - обращенной в сторону гаалоплазмы - поверхности мембраны аЭПС), (1) синтез гликогена, (3) синтез холестерина, (4) детоксикацию эндогенных и экзогенных веществ, (5) накопление ионов Са2+, (6) восстановление кариолеммы в телофазе митоза (эта функция оспаривается авторами, считающими, что кариолемма восстанавливается за счет мембранных пузырьков, на которые она ранее распалась). Помимо указанных основных функций, в некоторых типах клеток аЭПС выполняет ряд дополнительных - например, в мегакариоцитах (гигантских клетках костного мозга) ее элементы образуют демаркационные каналы, разделяющие формирующиеся тромбоциты.
Способность аЭПС к накоплению ионов Са2+ обусловлена наличием: (1)
кальциевого насоса в ее мембране, который обеспечивает транспорт этих ионов из гиалоплазмы внутрь цистерн аЭПС; (2) кальций-связывающих белков (кальсеквестрина в мышечных клетках, кальрети-
- 47 -
кулина - преимущественно в немьшечных и др.), которые в просвете цистерн образуют комплекс с ионами Са2+ и (3) кальциевых каналов в мембране аЭПС, которые осуществляют выведение Са2+ в гиалоплазму. Механизмы действия кальциевых каналов неодинаковы в клетках разных типов. Функция накопления ионов Са2+ особенно выражена в мышечных клетках, в которых специализированная аЭПС (именуемая саркоплазматической сетью) обеспечивает мышечное сокращение путем накопления и выделения значительных количеств ионов Са2+, связывающихся с особыми белками.
Обычно аЭПС в цитоплазме занимает меньший объем, чем грЭПС, однако она очень хорошо развита в клетках, синтезирующих стероиды, триглицериды и холестерин. Так, аЭПС занимает значительную часть объема цитоплазмы в клетках, которые активно продуцируют стероидные гормоны (клетки коркового вещества надпочечника, интерстициальные гландулоциты яичка (клетки Лейдига), клетки желтого тела яичника (лютеоциты) и др. Она также хорошо развита в клетках печени (гепатоцитах), где ее ферменты участвуют в процессах окисления, конъюгации и метилирования, которые обеспечивают нейтрализацию и детоксикацию ряда гормонов и вредных веществ (алкоголя, инсектицидов и др.).
Переходная (транзиторная) ЭПС - участок перехода грЭПС в аЭПС у формирующейся поверхности комплекса Гольджи. В области переходной ЭПС трубочки распадаются на отдельные фратенты, образующие окаймленные транспортные пузырьки, которые переносят материал из ЭПС в комплекс Гольджи (рис. 3-9).
Комплекс ГОЛЬДЖИ
Комплекс Гольджи - сложно организованная мембранная органелла, образованная тремя основными элементами - (1) стопкой уплощенных мешочков (цистерн), (2) пузырьками и (3) вакуолями, или секреторными пузырьками (см. рис. 3-1 и 3-9). Комплекс этих элементов называется диктиосомой (от греч. с11к1уоп - сеть); в некоторых клетках имеются множественные диктиосомы (до нескольких сотен). В специализированных секреторных клетках комплекс Гольджи располагается надъядерно под апикальной частью клетки, через которую происходит выделение секрета механизмом экзоцитоза. Нередко он лежит у ядра вблизи центриолей, в некоторых клетках его компоненты рассеяны по всей цитоплазме.
1. Цистерны имеют вид изогнутых дисков ("блюдец") диаметром 0.5-5 мкм и образуют стопку из 3-30 элементов, разделенных простран-
- 48 -
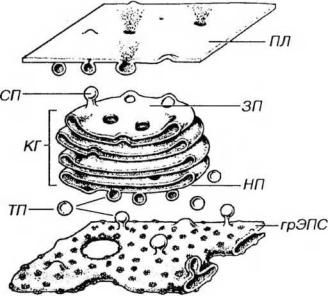
ством 15-30 нм; выпуклой стороной стопка обычно обращена к ядру, вогнутой - к плазмолемме. Каждая группа цистерн внутри стопки отличается особым составом ферментов, определяющим характер реакций процессинга белков. Периферические отделы цистерн несколько расширены, от них отщепляются пузырьки и вакуоли. Механизм, удерживающий стопку в виде единого образования, неизвестен. При наличии в клетке множественных диктиосом их цистерны связаны друг с другом системой анастомозирующих и ветвящихся трубочек.
Рис. 3-9. Синтетический аппарат клетки: грЭПС продуцирует белки, которые переносятся к незрелой поверхности (НП) комплекса Гольджи (КГ). От зрелой поверхности (ЗП) отделяются секреторные пузырьки (СП), содержимое которых выделяется за пределы клетки при слиянии мембраны СП с плазмолеммой (ПЛ)
2.Пузырьки - сферические окруженные мембраной элементы диаметром 40-80 нм с содержимым умеренной плотности; образуются путем отщепления от цистерн.
3.Вакуоли - крупные (диаметр - 0.1-1.0 мкм), окруженные мембраной сферические образования, отделяющиеся от цистерны на зрелой поверхности комплекса Гольджи (см. ниже) в некоторых железистых клетках. Они содержат секреторный продукт умеренной плотности, находящийся в процессе конденсации (конденсирующие вакуоли).
- 49 -
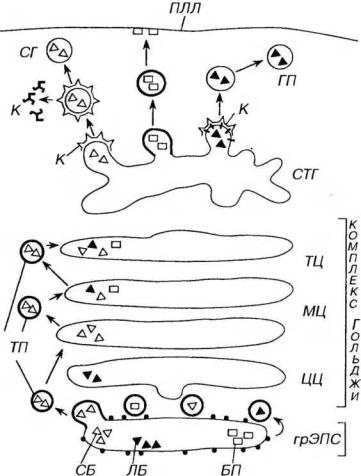
Рис. 3-10. Синтетический аппарат клетки (схема). ГрЭПС (синтез и начальный процессинг белков): СБ - секреторные белки, ЛБ - лизосомальные белки, БП - белки плазмолеммы; комплекс Гольджи (процессинг белков): ТП - транспортные пузырьки, ЦЦ - цисцистерны (комплекса Гольджи), МЦ - медиальные цистерны, ТЦ - трансцистерны, СТГ - сеть транс-Гольджи (сортировка белков), К - клатрин, СГ - секреторная гранула, ПЛ - первичная лизосома, ПЛЛ - плазмолемма, К - клатрин.
Полярность комплекса Гольджи. Комплекс Гольджи представляет собой поляризованную структуру, в которой выделяют две поверхности, обладающие структурными и функциональными различиями:
(а) цис- (от лат. а я - по эту сторону), незрелую, формирующуюся -
выпуклой формы, обращенную к ЭПС и связанную с системой мелких (транспортных) пузырьков, отщепляющихся от ЭПС;
- 50 -