Материал: Сердце
Конечно-систолический объем левого желудочка (КСО) – объем полости ЛЖ в конце систолы- 40-65 мл Ударный объем- объем крови, изгоняемой в систолу за одно сокращение. УО=КДО-КСО (70-100 мл)
Фракция выброса ЛЖ – отношение УО к КДО. Норма 50 -70 %
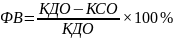
Определяется с помощью Эхокардиографии
КСР(конечный систолический размер)-22-40мм- размер в обхвате в момент систолы
КДР(конечный диастолический размер)(внутреннийц диаметр)-38-55мм- размер в обхвате в момент диастолы
Степень укорочения переднезаднего размера левого желудочка в систолу (%ΔS):
%ΔS=КДР-КСР/КДР •100% (норма 30-43%)
Используется для определения объема левого желудочка в конце систолы/диастолы, определения характера укорочения миокарда и наличии патологии(пр: инфаркт миокарда)
УО(ударный объем)-объем крови, поступающий в сосудистое русло при изгнании из желудочков
--разность между КДО и КСО= 60-80мл(в покое)
КСО(конечный систолический объем)-45-75мл
КДО(конечный диастолический объем)- 110-145мл
Вопрос№21. Физиологические свойства сердца (автоматия, проводимость, сократимость, возбудимость)
Возбудимость - это способность миокарда возбуждаться при действии раздражителя, сопровождающаяся изменениями физико – химических свойств ткани. Она меньше, чем у поперечно-полосатых скелетных мышц. Клетки рабочего миокарда имеют большую величину мембранного потенциала(70-90мв) и за счет этого реагируют только на сильное раздражение.
Проводимость – проводить возбуждение вследствие образования потенциала действия в клетках пейс-мейкерах
Сократимость – выполнение специфической функции—укорачиваться при возбуждении.
Автоматия- это способность сердца к самопроизвольным ритмическим сокращениям, возникающих в самом органе. Еще Аристотель писал, что в природе сердца имеется способность биться с самого начала жизни и до ее конца, не останавливаясь. В прошлом веке существовало 3 основных теории автоматии сердца.
Прохаска и Мюллер выдвинули нейрогенную теория, считая причиной его ритмических сокращений нервные импульсы.
Гаскелл и Энгельман предложили миогенную теорию, согласно которой импульсы возбуждения возникают в самой сердечной мышце. Существовала теория гормона сердца, который вырабатывается в нем и инициирует его сокращения. Автоматию сердца можно наблюдать на изолированном сердце по Штраубу. В 1902 году, применив такую методику профессор А.А.Кулябко впервые оживил человеческое сердце.
—в клетках атипического миокарда могут генерироваться нервные импульсы в области синоатриального узла
Клетки водители ритма первого порядка, или пейсмекеры:
-веретеновидная форма
-расположены группами
-окружены общей базальной мембраной
-низкая величина мембранного потенциала и высокая проницаемость для ионов Na и Ca
-низкая активность работы натрий-калиевого насоса, что обусловлено разностью концентрации Na и K
-с высокой скоростью идут обменные процессы→метаболиты не успевают выноситься и накапливаются в МК-жидкости
Механизм
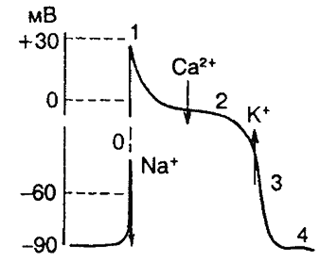
возникает в фазу диастолы и проявляется движением ионов Na внутрь клетки.
величина мембранного потенциала уменьшается и стремится к критическому уровню деполяризации – наступает медленная спонтанная диастолическая деполяризация, сопровождающаяся уменьшением заряда мембраны
В фазу быстрой деполяризации возникает открытие каналов для ионов Na и Ca, и они начинают свое движение внутрь клетки
заряд мембраны уменьшается до нуля и изменяется на противоположный, достигая +20–30 мВ.
Движение Na происходит до достижения электрохимического равновесия по ионам N a, затем начинается фаза плато.
В фазу плато продолжается поступление в клетку ионов Ca( В это время сердечная ткань невозбудима)
По достижении электрохимического равновесия по ионам Ca заканчивается фаза плато и наступает период реполяризации – возвращения заряда мембраны к исходному уровню.
Потенциал действия синоатриального узла отличается меньшей амплитудой и составляет ±70–90 мВ, а обычный потенциал ровняется ± 120–130 мВ.
В норме потенциалы возникают в синоатриальном узле за счет наличия клеток – водителей ритма первого порядка. Но другие отделы сердца в определенных условиях также способны генерировать нервный импульс. Это происходит при выключении синоатриального узла и при включении дополнительного раздражения.
При выключении из работы синоатриального узла наблюдается генерация нервных импульсов с частотой 50–60 раз в минуту в атриовентрикулярном узле – водители ритма второго порядка. При нарушении в атриовентрикулярном узле или при дополнительном раздражении возникает возбуждение в клетках пучка Гиса с частотой 30–40 раз в минуту – водитель ритма третьего порядка.
Вопрос№22. Условия, при которых проявляется свойство автоматии.
-отсутствие блокады проведения возбуждения
-отсутствие некроза ткани(ишемия)
-отстуствие перерезки проводимого волокна
-наличие нормальной трофики сердца
-наличие нормального восстановления мембранного потенциала
Вопрос№23.Методика изоляции сердца по Лангердорфу и Штраубу
Опыт Штрауба: канюля через аорту проведена в желудочек, благодаря питательному раствору (Раствор был назван в честь Сиднея Рингера, который в 1882–1885 годах установил, что в растворе для перфузии сердца лягушки должны содержаться соли натрия, калия и кальция в определённой пропорции, чтобы сердце продолжало биться в течение длительного времени) сердце может сокращаться в течение суток.
Оскар Лангендорф разработал первый препарат ex vivo по изучению изолированного сердца млекопитающих в 1895 году. В качестве перфузионной жидкости (перфузата) использовалось дефибрилированная кровь животных того же вида. В этом подходе коронарные сосуды перфузируются в обратном направлении (т.е. ретроградно) через аорту. Перфузии через коронарные сосуды было достаточно для обеспечения длительных сердечных сокращений. Однако, вследствие того, что нормальные пути циркуляции через желудочки не задействованы, эта модель не позволяет получать физиологически значимые данные по показателям "давление-объем", которые наблюдаются в целостном организме. В целом, препарат Лангендорфа обеспечивает только общую информацию по сердечной функции и дает данные, ограниченные динамикой в коронарных артериях.( раствор Рингера-Локка)
Вопрос№24. Виды кардиомиоцитов, их физиологическая характеристика
Кардиомиоцит – это вид миоцитов, представляющий собой основную структурно-функциональную единицу миокарда — миокардиальную клетку, ответственную за сократительную деятельность миокарда.
- окружены обильной сетью капилляров. Клетки проводящей системы, помимо капилляров, окружены вегетативными нервными окончаниями.
- соединяются друг с другом с помощью вставочных дисков (с помощью этих контактов за счет заполненных жидкостью каналов обеспечивается электрическое взаимодействие между кардиомиоцитами)
- окружены мембраной— сарколемма (в сарколемме имеется множество складок, выпячиваний и карманов, поверх нее имеется дополнительное рыхлое покрытие толщиной 50 нм, которое называется гликокаликсом. Гликокаликс связан с прилегающими к клетке капиллярами и участвует в обмене веществ между капиллярами и клеткой
- миофибриллы (содержат сократительные (миозин и актин) и регуляторные (тропомиозин и тропонин) белки.
Миозин образует толстые нити, или филаменты, актин — тонкие. Эти филаменты расположены параллельно друг другу, и каждая нить миозина окружена 6 нитями актина. Каждая нить актина, в свою очередь, окружена 6 нитями миозина. Диаметр толстых филаментов около 14 нм, длина — 1 500 нм, они находятся на расстоянии 20-30 нм друг от друга. Тонкие филаменты имеют диаметр примерно 7-8 нм.
- 2 или более ядер.
- на поверхности ядра имеется много углублений
- пластинчатый комплекс, содержащий углеводные и белковые остатки, липидные образования, гликоген
Классификация
по анатомическому положению
предсердные кардиомиоциты
желудочковые кардиомиоциты.
по функциональной деятельности
рабочие (сократительные) кардиомиоциты
- длина 100 мкм и диаметр – 15-20 мкм
- выполняют основную часть сократительной работы сердца
- основная масса миокарда (95-99%)
- отвечают за генерацию и распространение возбуждения, потенциалов действия по миокарду
- определяют частоту сокращений сердца и последовательность его возбуждения
- развит сократительный аппарат(большая часть сарколеммы)
проводящие (атипичные) кардиомиоциты
-больше и шире
-образуют узлы и пучки проводящей системы сердца
Р-клетки(Пейсмейкеры)
-генерируют электрические импульсы
-обеспечивают сердечный автоматизм (ритмическое сокращение сердца)
клетки Пуркинье
переходные кардиомиоциты (Т-клетки)
- между рабочими и проводящими
- проводят импульсы от проводящих кардиомиоцитов к рабочим.
секреторные (эндокринные)
- находятся в предсердиях (в особенности, правом)
- гормон (натрийуретический фактор (пептид) ПНФ (ПНП)) в предсердия, секреторные кардиомиоциты, таким образом, принимают участие в регуляции водно-электролитного баланса. вызывает усиленную потерю натрия и воды с мочой (натриурез и диурез), расширение сосудов, снижение артериального давления, угнетение секреции альдостерона, кортизола и вазопрессина
- отростчатая форма
- слабое развитие сократительного аппарата
- вблизи полюсов ядра находятся окруженные мембраной плотные
- гранулы диаметром 200-300 нм
- способностью к выработке ПНФ первоначально в ходе эмбрионального развития обладают все кардиомиоциты; в дальнейшем (уже после рождения) она резко падает в клетках желудочков, сохраняясь в предсердных кардиомиоцитах. При перегрузке сердечной мышцы способность к синтезу ПНФ может восстанавливаться в кардиомиоцитах желудочков
Гладкие миоциты не имеют поперечной исчерченности . Они содержат в своей центральной части по одному палочковидному ядру. Во многих клетках - большое количество гранулярной ЭПС. Здесь происходит синтез компонентов межклеточного вещества - протеогликанов, коллагена, эластина
Механизм сокращения гладкого миоцита
Фаза реполяризации
фаза медленной диастолической деполяризации - аналогична локальному (местному) потенциалу начинается сразу по завершении реполяризации и при достижении максимального диастолического потенциала. Самопроизвольную медленную диастолическую деполяризацию называют также пейсмекерным потенциалом клеток сердца, или предпотенциалом действия (снижается до критического уровня деполяризации, достигает его, что приводит к возникновению потенциала действия)
Компоненты сокращения
белки миокарда (актин и миозин)
«регуляторные» белки (тропонин, тропомиозин, кальмодулин)
Кальций
АТФ
В фазу диастолы актин и миозин диссоциированы. В начале деполяризации клеточной мембраны кардиомиоцита незначительное количество натрия поступает в клетку. Поступивший натрий вызывает цепь реакций, приводящих к освобождению кальция из системы эндоплазматического ретикулума и внутренней поверхности клеточной мембраны. поступает дополнительное количество кальция. Цитоплазматический кальций связывается с кальмодулином. Комплекс «кальмодулин-кальций» активизирует кининазу легких цепей миозина, что приводит к их фосфорилированию и утрате способности ингибировать взаимодействие актина с тяжелыми цепями миозина. Одновременно Цитоплазматический кальций, преимущественно освобождающийся из эндоплазматического ретикулума, соединяется с тропонином С. Этот процесс сопровождается конформационными изменениями тропонина и тропомиозина (способствует взаимодействию актина и миозина)
Поступление ионов Са2+. Под влиянием нервного импульса из внешней среды в клетку тем или иным способом (с помощью кавеол или через Са2+каналы) начинают поступать ионы Са2+.
Это происходит значительно медленней, чем выход Са2+ из цистерн в поперечнополосатых мышечных тканях. Поэтому сокращения гладкой мускулатуры развиваются не так быстро, как в тех тканях.
Фосфорилирование миозина. Еще одно отличие от тех же тканей состоит в том, что в гладких миоцитах ионы Са2+ влияют на состояние не тонких, а толстых миофиламентов. Причем это происходит опосредованным способом, а именно: ионы Са2+, связавшись с белком кальмодулином, активируют миозинкиназу (более точно — киназу легких цепей миозина), которая фосфорилирует молекулы миозина. В итоге миозин начинает объединяться в толстые миофиламенты, а последние — взаимодействовать с тонкими миофиламентами.
Взаимодействие миофиламентов. Толстые миофиламенты внедряются между тонкими — образуются временные миофибриллы. Далее, как обычно, миофиламенты перемещаются навстречу друг другу (за счет образования и разрыва мостиков и гидролиза АТФ). В результате плотные тельца сближаются, что и означает сокращение миоцита. В сокращенном состоянии гладкие миоциты могут пребывать достаточно долго без заметного утомления. Это объясняется тем, что часть миозиновых мостиков сохраняется и после дефосфорилирования миозина.
Выход из сокращения совершается тоже медленно. Его инициирует удаление ионов Са2+ из клетки Са2+насосами. После этого начинает преобладать активность миозинфосфатазы (точнее, фосфатазы легких цепей миозина). Происходит дефосфорилирование миозина. Еще какое-то время могут сохраняться некоторые миозиновые мостики.Тем не менее со временем толстые миофиламенты распадаются на фрагменты или даже на молекулы миозина. Клетка возвращается в расслабленное состояние.
механизм расслабления: Основной процесс, определяющий расслабление кардиомиоцитов, — удаление ионов Кальция из саркоплазмы, в результате чего концентрация Са2+ в ней уменьшается и становится ниже 10"7 моль/л. При этом комплексы Са2+ с тропонином С распадаются, тропомиозин смещается по отношению к актиновым филаментам и закрывает их активные центры — сокращение прекращается.
Вопрос№25. Сравнительная характеристика электрофизиологических особенностей рабочих и проводящих кардиомиоцитов, их ионные механизмы и значение.
ПД рабочего кардиомиоцита: