Дипломная (вкр): Определение влияния воды с измененным изотопным составом на функциональные особенности иммунокомпетентных клеток человека
Рисунок 2 - Фотография выделенных лимфоцитов,
сделанная на оптическом микроскопе БИОМЕД-6 Вар 3 при увеличении 1 : 1000
2.3 Инкубирование
лимфоцитов в воде с измененным изотопным составом и определение однонитевых
разрывов ДНК
Лимфоциты, изолированные из крови, инкубировались в течение 16 часов при комнатной температуре в физиологическом растворе, приготовленном на воде с пониженным содержанием дейтерия (40 ± 2 ppm) и обычной воде (150 ± 2ppm). Это необходимо для того, что бы выяснить, как влияет легкая вода на жизнеспособность клеток in vitro. Для выяснения влияния времени инкубирования на жизнеспособность лифоцитов, последние вымачивались в физрастворе, приготовленном на легкой воде в течение 3, 8, 16, 24 часов. Проведенные эксперименты показали, что при времени инкубирования 16 часов наблюдалась самая высокая эффективность репарации ДНК.
Для инкубирования две пробы лимфоцитов, выделенных из крови здоровых и больных ВПР ЧЛО доноров, замачивали в 1 мл физиологического раствора на легкой воде, а две другие аналогичные пробы замачивали в физиологическом растворе на обычной воде.
Для лизирования клеток был приготовлен 4,5М раствор мочевины. Мочевина является детергентом, используется в биологических и биохимических лабораториях, представляет собой мягкий реагент, который используются для разрушения мембраны клеток (лизиса клеток) и трансформации внутриклеточного материала в растворимую форму. Детергенты используются в основном для разрушения межбелковых, белково-липидных и межлипидных связей, денатурации белковых структур, предотвращения неспецифического связывания в иммунохимических анализах и кристаллизации белков. После инкубирования во все пробы было добавлено по 0,2 мл 4,5М раствора мочевины. Клетки лизировали в течение 10 мин при комнатной температуре. В этом случае происходит разрушение оболочки лимфоцитов, тем самом высвобождение молекулы ДНК.
Лизаты клеток в экспериментальных образцах подвергались щелочной обработке в течение 30 мин при 0 0С, а затем 60 минут при 15 0С. Для этого во все пробы добавлялось по 0,2 мл 0,1 % HCl. Далее образцы подвергались интенсивному встряхиванию на Вортексе. Этим достигалось расплетание молекул ДНК.
После щелочной обработки лизатов в образцы добавлялся раствор бромистого этидия. Это химическое вещество из группы фенантридинов (3,8-диамино-6-этил-5-фенилфенантридиумбромид), широко применяемое для флуорисцентной маркировки ДНК. При облучении бромистый этидий флюоресцирует оранжевым цветом. Является интеркалирующим агентом, т. е. веществом, способным встраиваться между основаниями ДНК, его интеркаляция сопровождается раскручиванием суперспирализованных участков ДНК. Используется в качестве красителя для детектирования двухцепочечных молекул ДНК и РНК.
Интенсивность флюоресценции полученных образцов определялась на спектрофлуориметре Флюорат-02-Панорама в диапазоне длин волн 540-740нм.
Количество однонитевых разрывов ДНК оценивалось по отношению величин флюоресценции контрольных и экспериментальных образцов. Для снятия фоновой флуорисценции подготовили образец, содержащий все компоненты, как в экспериментальных образцах и в той же концентрации, но без молекул ДНК. Помещаем образец в кюветное отделение. На компьютере открываем программу «Панорама», в ней выбираем режим флуориметрии. Затем задаем длину волны возбуждения - 540 нм, спектральный диапазон регистрации 540-740 нм. Далее нажимаем кнопку старт. Измерение флуоресценции в данном диапазоне идет около 2 минут для каждого образца. Полученные графики экспортируем непосредственно в электронную таблицу Excel для дальнейшего анализа результатов.
В ряде экспериментов использовался
рекомбинантный человеческий фактор некроза опухоли hTNFα.
При
замачивании лимфоцитов здоровых доноров в физиологическом растворе на обычной
воде и воде с пониженным содержанием дейтерия в среде создавалась концентрация
hTNFα
равная
10 нг/мл.
2.4 Изучение влияния
воды с измененным изотопным составом на состояние ДНК лимфоцитов
Для изучения влияния воды с пониженным содержанием дейтерия на жизнеспособность иммунокомпетентных клеток in vitro, лимфоциты, изолированные из крови здоровых доноров и больных с ВПР ЧЛО, инкубировались в физрастворах, приготовленных на воде с природным содержанием дейтерия (содержание D 150 ± 2 ppm) и на воде с измененным изотопным составом (содержание D 40 ± 2 ppm). После лизирования клеток и соответствующей обработки растворов получены спектры флюоресценции.
Рисунок 4 - Интенсивность
флюоресценции раствора бромистого этидия с лизатами лимфоцитов крови здоровых
доноров. (Инкубирование в физрастворах, приготовленных на обычной
дистиллированной воде (содержание D 150 ppm) и воде с измененным изотопным
составом (содержание D 40 ppm). Время инкубирования 16 час, t = 24 0С.)
На рисунке 4 представлены снятые спектры флюоресценции для здоровых доноров. При замачивании в физрастворе на обычной и легкой воде величина флюоресценции от фона не отличается, что говорит об отсутствии однонитевых разрывов ДНК лимфоцитов.
На рисунке 5 представлены снятые спектры флюоресценции для больных с ВПР ЧЛО доноров, ДНК которых имеет множество разрывов. Из рисунка видно, что при инкубировании лимфоцитов физрастворе на легкой воде флюоресценция образцов выше, т. е. количество однонитевых разрывов меньше, чем при инкубировании в физрастворе на обычной воде.
Таким образом, в ходе выполнения
эксперимента показано, что вода с пониженным содержанием дейтерия является либо
ингибитором апоптоза иммунокомпетентных клеток, либо активирует их
ДНК-репарирующие системы.
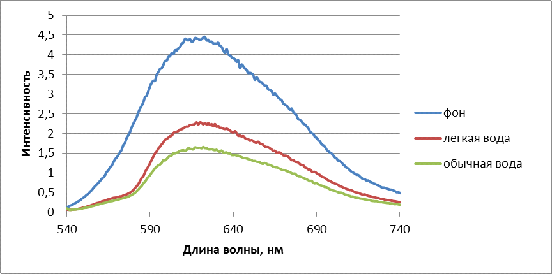
Рисунок 5 - Интенсивность
флюоресценции раствора бромистого этидия с лизатами лимфоцитов крови больных с
ВПР ЧЛО. (Инкубирование в физрастворах, приготовленных на обычной
дистиллированной воде (содержание D 150 ppm) и воде с
измененным изотопным составом (содержание D 40 ppm). Время
инкубирования 16 час, t = 24 0С.)
Для выяснения данного предположения
были проведены эксперименты с использованием рекомбинантного человеческого
фактора некроза опухолей hTNFα. Как известно [36], hTNFα является
провоспалительным цитокином, который инициирует апоптоз как в лимфоцитах, так и
в нейтрофилах. В экспериментах в среде создавалась концентрация hTNFα, равная 10
нг/мл, которая гарантированно инициирует апоптоз лимфоцитов. Клетки, выделенные
из крови здоровых доноров, инкубировались в физрастворе, приготовленном на воде
с пониженным содержанием дейтерия, в присутствии hTNFα при комнатной
температуре. Для контроля лимфоциты инкубировались в физрастворе,
приготовленном на обычной дистиллированной воде, также в присутствии и в
отсутствии hTNFα. По
истечении времени инкубации проводили измерения количества однонитевых разрывов
ДНК. На рисунке 6 приведены соответствующие спектры флюоресценции растворов
лизатов клеток с бромистым этидием. Из рисунка видно, что при использовании
обычной воды действие фактора некроза hTNFα приводит к прогрессивному
накоплению однонитевых разрывов ДНК в клетках - интенсивнось флюоресценции
снижалась. При замачивании на легкой воде разрывов ДНК не появлялось, что
говорит защищающем действии легкой воды.
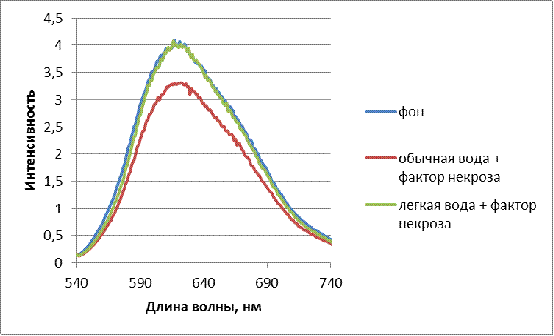
Рисунок 6 - Интенсивность
флюоресценции раствора бромистого этидия с лизатами лимфоцитов крови здоровых
доноров. (Инкубирование в физрастворах с 10 нг/мл hTNFα,
приготовленных на обычной дистиллированной воде (содержание D 150 ppm) и воде с
измененным изотопным составом (содержание D 40 ppm). Время
инкубирования 16 час, t=24 0С.)
На рисунке 7 представлена диаграмма
процентного содержания однонитевых разрывов ДНК в норме, при добавлении фактора
некроза и больных с ВПР ЧЛО при инкубировании в физиологическом растворе на
легкой и обычной воде. Обнаружено, что инкубация лимфоцитов больных доноров в
физрастворе, приготовленном на воде с содержанием дейтерия 40 ppm за 16 часов
уменьшает количество однонитевых разрывов ДНК по сравнению с контролем с 59 -
62 % до 36 - 40 %. Увеличение времени инкубирования до 24 час существенно не влияло
на полученные значения.
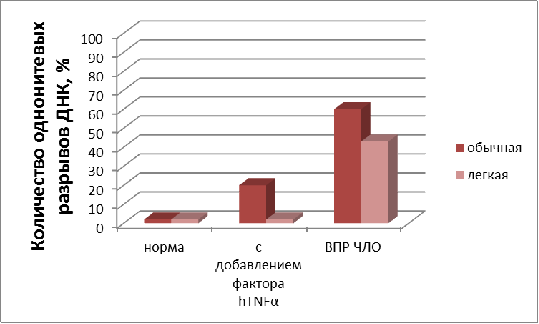
Рисунок 7 - Влияние воды с
измененным изотопным составом на количество однонитевых разрывов лимфоцитов в
норме, с добавлением в инкубационную среду 10 нг/мл hTNFα и у больных
ВПР ЧЛО. (Инкубирование в физрастворах, приготовленных на обычной
дистиллированной воде (содержание D 150 ppm) и воде с
измененным изотопным составом (содержание D 40 ppm). Время
инкубирования 16 час, t = 24 0С.)
При аналогичной обработке лимфоцитов, полученных из крови здоровых доноров, количество однонитевых разрывов ДНК было существенно меньше, мало зависело от среды инкубирования и составляло 2,5 ± 0,2 %. Это свидетельствует, о том, ДНК лимфоцитов здоровых доноров менее подвержены повреждениям по сравнению с ДНК лимфоцитов больных с ВПР ЧЛО, в которых наблюдаются мутации генов.
При инкубировании лимфоцитов здоровых доноров с добавлением фактора некроза опухоли в физрастворе на обычной воде, количество разрывов ДНК увеличивалось с 2,5 ± 0,2%до 23 - 25 %, а при замачивании на легкой воде, однонитевых разрывов не наблюдалось.
При использовании воды с содержанием дейтерия 40 ppm количество однонитевых разрывов в лимфоцитах достигало максимума через 2 часа инкубации, а затем резко снижалось. Такое поведение клеток в воде с пониженным содержанием дейтерия, возможно, связано с тем, что наряду с возникновением однонитевых разрывов в клетках активируются системы ее репарации, ликвидирующие эти разрывы. Появлению однонитевых разрывов ДНК предшествует ее фрагментация, которая наблюдается при вхождении клеток в стадию апоптоза. Вода с пониженным содержанием дейтерия, по-видимому, делает обратимым этот процесс.
Повреждения ДНК могут быть ликвидированы с помощью различных механизмов. Основные репаративные механизмы вырезания (BER) и механизмы репарации нуклеотидов (NER) позволяют удалить поврежденные и модифицированные нуклеотиды. В частности, NER работает главным образом на спирали ДНК, препятствуя транскрипции поврежденных участков, в то время как BER удаляет одиночные нуклеотиды, модифицированные с помощью метилирования, алкилирования, дезаминирования, или окисления.
Одно- и двунитевые разрывы ДНК являются частыми событиями в эукариотических клетках: либо при физиологических условиях в лимфоцитах, либо при патологических процессах, при окислительном стрессе, ионизирующем излучении, нарушении функционирования ферментов клеточного ядра.
Адаптация клеток к пониженным концентрациям дейтерия в среде может способствовать увеличению функциональной активности систем клетки, связанных со значительными траффиками ионов водорода и последующей активацией системы ДНК-репарации, связанной с большими затратами энергии.
Среда с пониженным содержанием
дейтерия создает более благоприятные условия функционирования клеток иммунной
системы, о чем свидетельствует большая сохранность лимфоцитов при инкубации в
подобной среде.
ЗАКЛЮЧЕНИЕ
Основные результаты дипломной работы состоят в следующем:
изучена литература, посвященная воздействию воды с измененным изотопным составом на живые системы;
проведены исследования влияния воды с измененным изотопным составом на состояние ДНК лимфоцитов в норме и патологии;
экспериментально показано, что вода с пониженным содержанием дейтерия уменьшала количество однонитевых разрывов ДНК лимфоцитов больных с ВПР ЧЛО от 60% до 43%, так же легкая вода защищала ДНК лимфоцитов от повреждения, наносимого фактором hTNFα;
проверен и подтвержден факт активизации репаративной системы лимфоцитов при их инкубации в воде с пониженным содержанием дейтерия;
показано, что вода с пониженным содержанием дейтерия предотвращает апоптоз (массовую гибель) иммунокомпетентных клеток, что обеспечивает их большую сохранность при воздействии повреждающих факторов (некроз, ионизирующие излучения, токсины и др.);
впервые получены данные о влиянии воды с пониженным содержанием дейтерия (40 ppm) in vitro на некоторые функциональные особенности иммунокомпетентных клеток в норме и патологии.
Это открывает перспективы для использования такой воды при различных видах ВПР с целью предотвращения неконтролируемой массовой гибели иммунокомпетентных клеток.
Результаты дипломной работы изложены в следующих публикациях:
Текуцкая Е.Е., Барышев М.Г. Сингур З.Н. Хачатрян А.Ш. Захарова Н.А. «Изучение воздействия низкочастотного электромагнитного поля на молекулы ДНК в водных растворах» в коллективной монографии // Современные проблемы физики, биофизики и инфокоммуникационных технологий»;
Текуцкая Е.Е., Барышев М. Г., Джимак С. С., Васильев Ю.А., Захарова Н.В. Влияние воды с пониженным содержанием дейтерия на иммунокомпетентные клетки человека в норме и патологии.» // Бюллетень экспериментальной биологии и медицины:
Захарова Н.А., Текуцкая Е.Е.,
Храмцова А.А., «Влияние воды с измененным изотопным составом на лимфоциты
человека в норме и паталогии» // Материалы научно-практической конференции
студентов и аспирантов.
СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ
1 Петровский. Б. В. Краткая медицинская энциклопедия / Б. В. Петровский. - М.: Советская энциклопедия, 1989. - 670 с.
Галактионов В. Г. Эволюционная иммунология / В. Г. Галактинов - М.: ИКЦ «Академкнига», 2005. - 408 с.
Воробьев А. В. Микробиология / А. В. Воробьев, А. С. Быков, Е. П. Пашков - М.: Медицина, 1998. - 336 с.
Ярилин А. А. Основы иммунологии / А. А. Ярилин - М.: Медицина, 1999. - 752 с.
Молекулярные механизмы апоптоза / В. К. Кухта, Н. В. Морозкина, Е. В. Богатырева, и др. // БМЖ. - 2004. - № 1. - С. 1-8.
6 Brune B. Nitric oxide: NO apoptosis or turning it ON? / Brune B. // Cell Death Differ. - 2003. - Vol. 10. - № 8. - P. 864-869.
Ghobrial I. M. Targeting apoptosis pathways in cancer therapy / I. M. Chobrial, T. E. Witzig, A. A. Adjei / CA Cancer J. Clin. - 2005. - Vol. 55. - P. 178-194.
Szende B. The occurrence and significance of apoptosis in tumors / B. Szende // Magy Onkol. - 2004. - Vol. 48. - № 3. - P. 215-219.
Vermeulen K. Apoptosis: mechanisms and relevance in cancer / K. Vermeulen, D. R. Van Bockstaele, Z. N. Berneman // Ann. Hematol. - 2005. - Vol. 84. - P. 627-639.
Широкова А. В. Апоптоз. Сигнальные пути и изменение ионного и водного баланса клетки / А. В. Широкова // Цитология. - 2007. - Т. 49. - № 5. - С. 385-394.
Манских В. Н. Морфологические методы верификации и количественной оценки апоптоза / В. Н. Манских // Бюллетень сибирской медицины. - 2004. - № 1. - C. 63-70.
Сингер М. Гены и геномы / М. Сингер, П. Берг - М.: Мир, 1998. - 373 с.
Стент Г. Молекулярная генетика / Г. Стент, Р. Кэлиндар - М: Мир, 1981. - 646 с.
Глик Б. Молекулярная биотехнология. Принципы и применение / Б. Глик, Дж. Пастернак - М.: Мир, 2002. - 592 с.
Курчанов Н. А. Генетика человека с основами общей генетики / Н. А. Курчанов - СПб.: СпецЛит, 2009. - 191 с.
Тронов В. А. Остаточные повреждения ДНК в периферических лимфоцитах после тотального облучения человека / В. А. Тронов, Е. В. Гринько, М. В. Кончаловский // Медицинская радиология. - 1993. - Т. 38. - № 2. - С. 38-41.
17 Nataraiau A. I. DNA repair and chromosomal alterations / A. I. Nataraiau, K. Palilti // Mutdtioh Research. - 2008. - Vol. 657. - № 1. - P. 3-7.
Kolodner R. D. Coupling distain sates in DNA during DNA mis-match repair / R. D. Kolodner, M. M. Mendillo, C. D. Putnam // Proceedings of the Natural Academy of Sciences of the United Slates of America. - 2007. - Vol. 104. - № 32. - P. 12953 - 12955.
Hoctimakers J. H. Genome maintenance mechanisms for reverting cancer / J. H. Hoctimakers // Nature. - 2001. - Vol. 411. - № 6835. - P. 366-374.
Lifter M. R The mechanism of double-strand DNA break repair by the nonhomologous DNA end joining pathway / M. R. Lifter // Annual Review of Biochemastry. - 2010. - Vol. 79. - № 6. - P. 181-211.
Shibata A. Factors determining DNA double strand break repair pathway choice in G12 phase / A. Shibata, S. Conrad // EMBO Journal. - 2011. - Vol. 30. - № 6. - P. 1079-1092.
22 Антонченко В. Я. Основы физики воды / В. Я. Антонченко, А. С. Давыдов, В. В. Ильин - Киев: Наук. Думка, 1991. - 672 с.
Баранова В. Ю. Изотопы: свойства, получение, применение / В. Ю. Баранова - М.: ИздАТ, 2000. - 703 с.
Бармин М. И. Экология воды / М. И. Бармин, П. Н. Темнов - M.: Комильфо, 2010. - 237 с.
Мухачев В. М. Живая вода / В. М. Мухачев. - М.: Наука, 1975. - 143 c.
26 Bild W. Research concerning the radioprotective and im-munostimulating effects of deuterium-depleted water / W. Bild, I. Stefanescu, I. Haulica // Rom J Physiol. - 1999. - № 36. - P. 205-218.
Doina P. M. Variation of the deuterium concentration in rats» blood after deuterium depleted water administration and intoxication with cadmium / P. M. Doina, L. Olariu, S. Cun // Bulletin UASVM Veterinary Medicine. - 2008. - Vol. 65. - № 1. - P. 418-423.
28 Мосин О. В. Изучение биологических эффектов тяжелой воды в биологических системах на примере клеток прокариот и эукариот / О. В. Мосин, И. Игнатов // Биомедицина. - 2012. - №3. - С. 35-50.
Лобышев В. Н. Изотопные эффекты D2О в биологических системах. / В. Н. Лобышев, Л. П. Калиниченко - М.: Наука, 1978. - 215 с.