Материал: Лекция 8
Онкогенные ДНК-содержащие вирусы
ДНК-содержащие вирусы , способные индуцировать опухоли
24
ДНК-содержащие онкогенные вирусы
Тип вируса |
Окогенность у |
Онкогенность у |
|
животных |
человека |
||
|
|||
|
|
|
|
Аденовирусы |
Да. Через ранние гены |
Нет |
|
|
Е1А и Е1В |
|
|
|
|
|
|
Герпесвирусы |
Да. Механизм |
Да. Возможно через |
|
|
неизвестен |
гены группы EBNA и |
|
|
|
LMP |
|
|
|
|
|
Вирус гепатита В |
Да. Через |
Да. Через |
|
|
трансактивирующий |
трансактивирующий |
|
|
ген Х |
ген Х |
|
|
|
|
|
Вирус группы полиомы |
Да. Через гены, |
Нет |
|
|
кодирующиеT-антиген |
|
|
|
|
|
|
Вирусы папиллом |
Да. Механизм сходен с |
Да. Через |
|
|
вирусом папилломы |
трансформирующие |
|
|
человека |
механизмы Е6 и Е7 |
|
|
|
|
26
Онкогенные ДНК-содержащие вирусы
•Группа вирусов с различными структурой, организацией генома и стратегией репликации.
•Индуцируют опухоли у природных хозяев
–Вирусы папилломы
–Вирус Эпштейна-Барр,
–Герпесвирус ассоциированный с саркомой Капоши
–Гепатит В
•Индуцируют опухоли в экспериментальных системах:
–Аденовирусы
–Полиомавирусы ,Обезьяний вирус -40 (SV40)
Онкогены ДНК-содержащих вирусов - истинные вирусные онкогены.
25
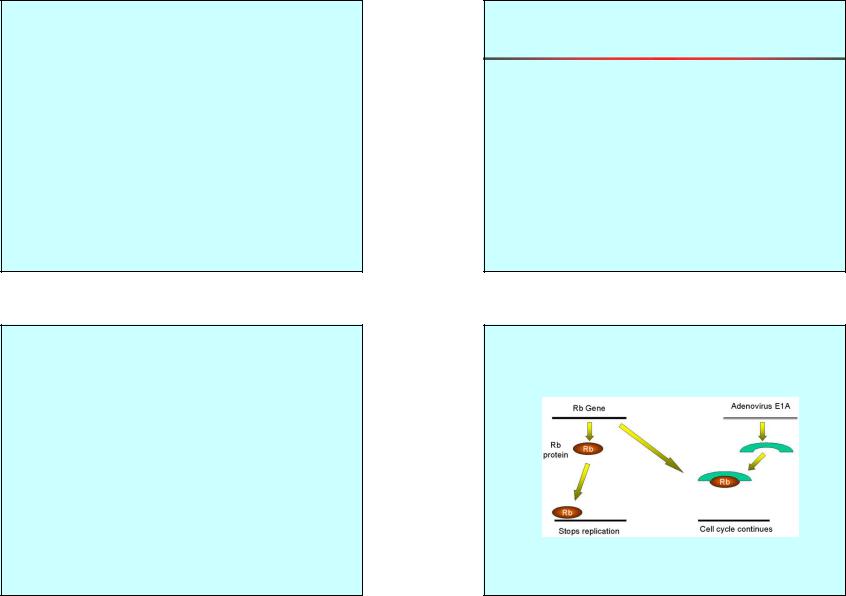
Онкоген аденовируса инактивирует супрессор опухоли.
27
6
Онкоген инактивирует супрессор опухоли.
Е6
Т-антиген
Polyoma
28
ДНК-содержащие онкогенные вирусы
36
Онкогенные вирусы
|
Ретровирусы |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
ДНК – |
содержащие вирусы |
|
|||||||||||
Инсерционный |
Вносят |
|
|
|
||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
мутагенез |
вирусные |
|
|
|
|
|
Вносят |
|
|
|
|
|||||||
|
|
|
онкогены |
|
Инсерционный |
вирусные |
||||||||||||
|
|
|
|
|
|
|
|
онкогены |
||||||||||
|
|
|
|
|
|
|
|
|
мутагенез |
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
TAX |
|
|
|
|
|
|
|
|
|
|
|
E7,E1A |
|
|
|
|
|
|
|
|
|
E6,LMP1 |
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
E1B |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
T-SV40 |
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
Активзированные |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
клеточные |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
протоонкогены |
|
|
|
|
|
|
|
|
|
|
Активзированные |
|
|
Изменение |
|
Подавление |
|
Подавление |
|||||||||||
|
|
|
|
|
|
|
||||||||||||
клеточные |
|
транскрипции |
|
|
|
|
активности |
|
активности |
|||||||||
протоонкогены |
|
клеточных белков |
|
|
|
|
р53 |
|
|
|
pRb,p105,p130 |
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Стимуляция размножения и миграции клеток, Отмена негативного контроля пролиферации Подавление апоптоза, генетическая нестабильность
35
•ДНК-содержащие онкогенные вирусы являются модельной системой для исследования молекулярной биологии эукариотической клетки:
репликация генома и экспрессия генов ДНК- содержащих вирусов идет с использованием механизмов и ферментативного аппарата клетки.
37
7

ПАПОВАВИРУСЫ
АДЕНОВИРУСЫ
•Наиболее хорошо изученные ДНК-содержащие вирусы, индуцирующие опухоли.
•В природных хозяевах, в пермиссивных клетках, инфекционный процесс приводит к репликации вируса, лизису зараженных клеток и освобождению вирусного потомства
•Используют один и тот же механизм
трансформации клеток и являются экспериментальными моделями для изучения причин образования опухолей
•Не связанны с онкологическими заболеваниями человека.
Группа I
38
сем. Polyomaviridae род Polyomavirus
Simian Virus (SV40)
Обезьяний вирус -40
Реплицируются в ядре |
Simian Vacuolating Agent 40 |
42
ПАПОВАВИРУСЫ
PApillomaviridae
POlyomaviridae
Simian Vacuolating Agent
( SV-40 - Polyomavirus genus)
PAPOVA
39
Обезьяний вирус - 40 (SV-40)
•Икосаэдр, 72 капсомера (12 пентомеров, 60 гексамеров)
•Три структурных белка VP1 (40 kD),VP2 (38kD), VP3 (27kD);
•Замкнутая кольцевая ds ДНК 5243 пн связана с клеточными гистонами H2a и b, H3,H4 (минихромосома)
Т=7
•Неструктурные регуляторные белки большой Т-антиген
(82 kD) и малый t-антиген (17kD)
43
8
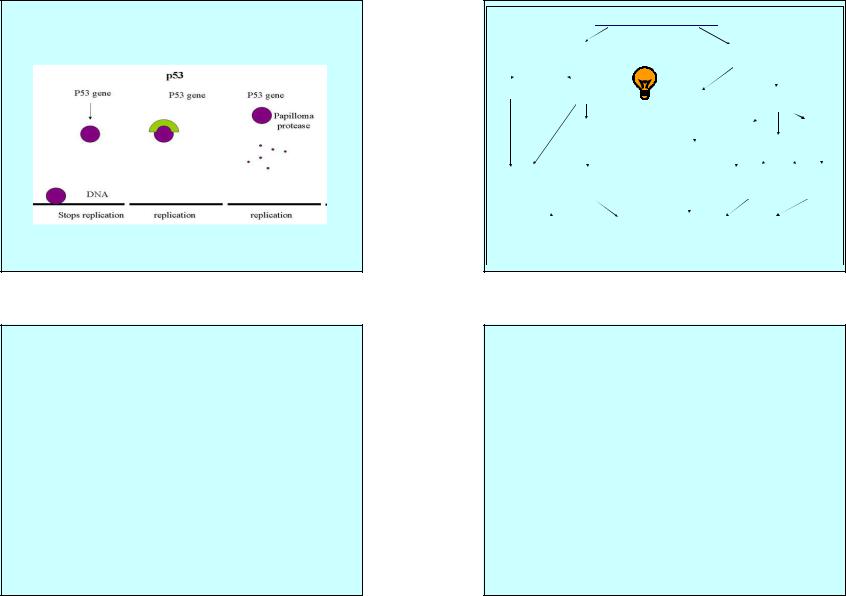
Ответ клетки на заражение SV40
|
Инфекция |
продуктивная |
непродуктивная |
(пермиссивные клетки) |
(непермиссивные клетки) |
Клетки обезьяны |
Клетки грызунов |
лизис клеток |
трансформация |
|
клеток |
44
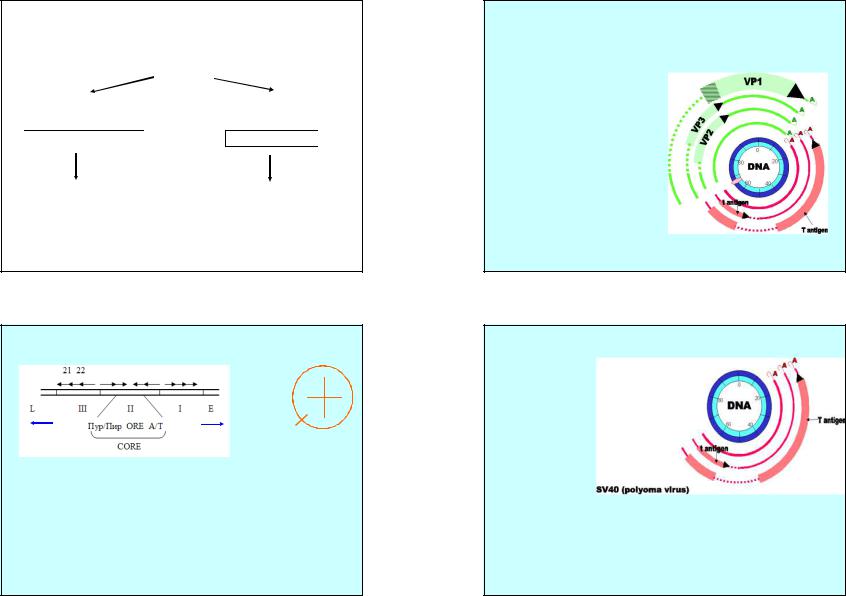
Структура Ori SV40
0
0,75
0,25
A/T |
Пур/Пир |
0,5 |
Ori 0,67
(I)пентануклеотидные последовательности GAGGC , организованные как инвертированный повтор и узнаваемые T- антигеном.
(II) CORE длиной 64 пн (Core Ori Recognition Element) состоит из центрального участка, с которым связывается T-антиген, и фланкирован c одной стороны А/Т богатым участком, с другой - Пур/Пир последовательностью.
(III) участке находятся повторы 21,22 п.н. – участок переключения с ранней на позднюю стадию транскрипции
Общая длина Ori со всеми прилегающими участками – 170 п.н.
47
Процесс экспрессии вирусных белков делится на две фазы: раннюю и позднюю
•РАННИЕ гены кодируют ферменты и регуляторные белки, необходимые для начала репликации
•ПОЗДНИЕ гены кодируют структурные белки, необходимые для сборки и созревания вируса.
46
Ранняя транскрипция генов
Клеточная РНК- полимераза II
Посттранскрипцион- ная модификация
Образуется первичный транскрипт и подвергается альтернативному сплайсингу, в результате которого образуются мРНК для малого t- и большого Т-антигена ( белки имеют общий N-конец и различные С- концы полипептидной цепи).
мРНК транслируется в цитоплазме.
48
9
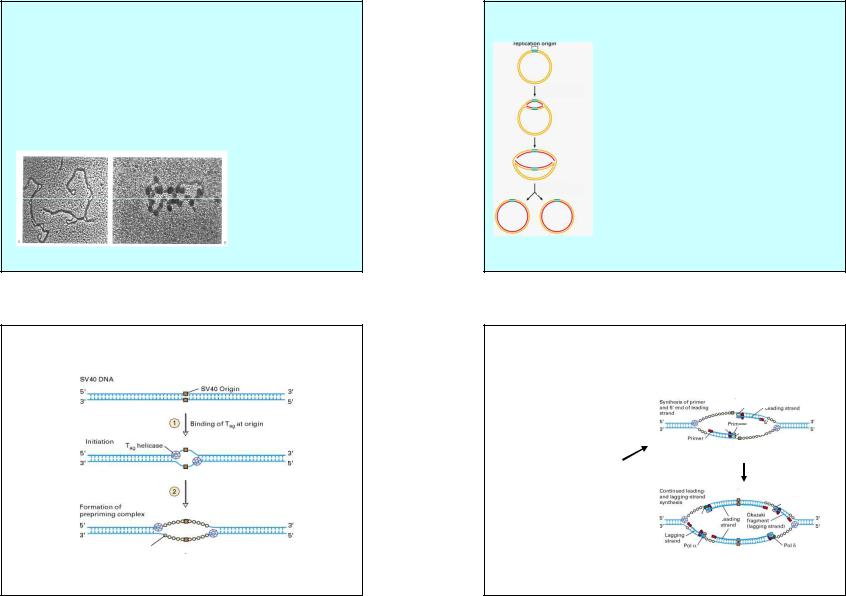
Поздняя фаза литического цикла начинается с репликации вирусного генома
•Вирусный геном образует комплекс с клеточными гистонами – минихромосому, неотличимую от клеточного хроматина.
•Вирус использует клеточный аппарат репликации ДНК.
•Механизм репликации ДНК SV40 сходен с репликацией клеточной ДНК
ДНК SV40 |
минирохромосома |
49 |
|
|
Репликация ДНК SV-40
SSB
50a
Репликация ДНК SV-40
•Репликация ДНК по схеме Кернса происходит в ядре
•Т-антиген необходим
•Т-антиген изменяет локальную структуру Ori
•Т-антиген взаимодействует с праймазой, образуется нуклеотидная затравка
•Клеточная ДНК-полимераза узнает Ori –
участок начала репликации, только, если с ним связан Т-антиген
50
Репликация ДНК SV-40
DNA-Pol
DNA-Pol
Т-антиген
ПРАЙМАЗА
SSB
ДНК-ПОЛИМЕРАЗА II ХЕЛИКАЗА (?)
DNA-Pol
DNA-Pol
DNA-Pol |
|
DNA-Pol |
50b
10