Материал: Изучение структуры генетического материала человека
Изучение структуры генетического материала человека
Содержание
Введение
. Строение ДНК
. Природа генетического кода
.1 Триплетность
.2 Смысл кодонов
.3 Специфичность
.4 Вырожденность
.5 Линейность записи информации
.6 Универсальность
.7 Колинеарность гена и продукта
. Организация генетического материала в хромосомах человека
Заключение
Использованная
литература
Введение
В соответствии с современными представлениями все процессы клеточного метаболизма, лежащие в основе жизнедеятельности организма, находятся под контролем генетической программы, которая содержится в структуре молекул нуклеиновых кислот и определяет, в первую очередь, особенности строения всех белков, синтезируемых в клетке.
Эта программа закодирована в виде специфического чередования нуклеотидов молекул дезоксирибонуклеиновой кислоты (ДНК), находящихся в хромосомах вирусов, нуклеоидов бактерий и ядер эукариот, а также в экстрахромосомных (цитоплазматических) структурах клеток (плазмиды бактерий, ДНК митохондрий и хлоропластов).
Молекулы рибонуклеиновых кислот (РНК) обеспечивают реализацию (декодирование) генетической информации, содержащейся в молекулах ДНК.
У РНК-содержащих вирусов первичным генетическим материалом является РНК.
Первые прямые доказательства роли ДНК как хранителя и переносчика генетической информации получил О. Эвери с сотрудниками (О. A very et al., 1944) в экспериментах по трансформации бактерий. Эти авторы показали, что проникновение молекул очищенной ДНК, выделенной из вирулентных пневмококков, вызывающих заболевание и гибель зараженных мышей, в клетки авирулентного штамма этих бактерий может сопровождаться превращением (трансформацией) последних в вирулентную форму.
Наиболее убедительные современные доказательства генетической роли ДНК связаны с разработкой методов генной инженерии, позволяющих искусственно конструировать гибридные (рекомбинантные) молекулы ДНК, кодирующие синтез генных продуктов (белков), интересующих исследователя.
Благодаря современным методам молекулярной
биологии и молекулярной генетики были получены сведения об особенностях
строения и функционирования генетического материала многих организмов,
находящихся на разных уровнях организации живой материи.
1. Строение ДНК
Молекула нуклеиновой кислоты представляет собой полимер (полинуклеотид), состоящий из последовательно соединенных друг с другом мономеров (нуклеотидов).
В свою очередь, каждый нуклеотид представляет собой соединение, в котором присутствуют три различные молекулы: остаток фосфорной кислоты (фосфат), углевод (пентоза) и азотистое основание (пуриновое либо пиримидиновое). Принципиальная схема строения нуклеотида приводится на рис. 1.
Следует отметить, что нуклеотиды молекул ДНК (дезоксирибонуклеотиды) содержат углевод дезоксирибозу и одно из четырех азотистых оснований - аденин (сокращенно обозначается символом А), гуанин (Г), тимин (Т) и цитозин (Ц), первые два из которых являются производными пурина, а два последних - производными пиримидина.
В состав нуклеотидов РНК (рибонуклеотидов) входит другая пентоза (рибоза) и также одно из четырех азотистых оснований - аденин, гуанин, урацил (У) и цитозин (вместо тимина здесь включается пиримидиновое основание урацил). Поскольку в составе молекулы пентозы имеется 5 атомов углерода, то каждый из них можно пронумеровать индексом от Г до 5' (см. рис. 1). В каждом нуклеотиде присоединение азотистого основания происходит к первому углеродному атому (Г) пентозы с помощью TV-гликозидной связи.
Соединение, состоящее из углевода (пентозы) и азотистого основания, называется нуклеозидом (рис. 2).
Формирование линейной полинуклеотидной цепочки
(первичной структуры молекулы нуклеиновой кислоты) происходит при соединении
пентозы одного нуклеотида с фосфатом другого нуклеотида путем образования
фосфодиэфирной связи (рис. 3). При этом в зависимости от порядкового номера
углеродного атома (3'либо 5') концевой молекулы пентозы, участвующего в
образовании фосфодиэфирной связи с фосфатом, такая цепочка имеет маркированный
3'-конец и 5'-конец.
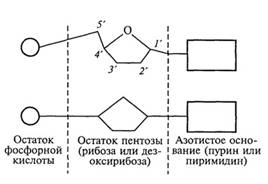
Рис. 1. Два варианта схематического изображения
строения нуклеотида: 1' 5' атомы углерода в молекуле углеводорода (пентозы)
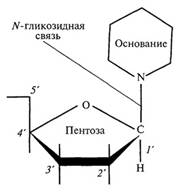
Рис. 2. Строение нуклеозида
Расшифровка генетической информации,
содержащейся в молекулах ДНК, оказалась возможной лишь после установления
структурных особенностей этих молекул в работах Дж. Уотсона и Ф. Крика (I.
Watson, F. Crick, 1953). Предпосылкой для создания их модели молекулы ДНК
послужили результаты биохимических исследований Э.Чаргаффа (Е. Chargaff, 1950),
а также данные рентгеноструктурного анализа. При изучении препаратов ДНК, полученных
из клеток организмов разных видов, Э.Чаргафф установил правило эквивалентности,
согласно которому почти в любом образце ДНК молярное содержание аденина
практически равно молярному содержанию тимина, а содержание гуанина равно
содержанию цитозина, т.е. А = Т и Г = Ц. Вместе с тем, соотношение пар А-Т и Г
-Ц (показатель (А + Т)/(Г + Ц)) имело значительные колебания при сравнении
образцов ДНК из организмов разных видов. Согласно модели Уотсона-Крика молекула
ДНК состоит из двух полинуклеотидных цепочек (нитей, тяжей), соединенных друг с
другом с помощью поперечных водородных связей между азотистыми основаниями по
комплементарному принципу (аденин одной цепочки соединен двумя водородными
связями с тимином противоположной цепочки, а гуанин и цитозин разных цепочек
соединены друг с другом тремя водородными связями).
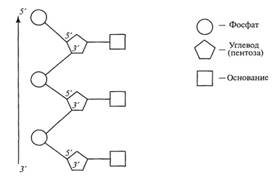
Рис. 3. Образование полинуклеотида
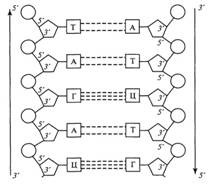
Рис. 4. Схематическое изображение первичной
структуры фрагмента двух-цепочечной молекулы ДНК: А аденин; Г гуанин; Т тимин;
Ц цитозин
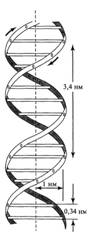
Рис. 5. Модель вторичной структуры ДНК Уотсона
Крика
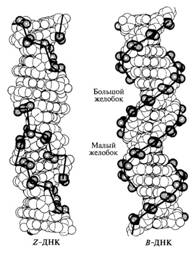
Рис. 6. Пространственные модели Z-формы и
5-формы ДНК
Таблица 1 Свойства различных форм двойных спиралей ДНК
|
Свойства |
Формы спиралей |
|||
|
|
А |
B |
С |
Z |
|
Направление скрученности |
Направо |
Направо |
Направо |
Налево |
|
Расстояние между соседними парами оснований (нм) |
0,23 |
0,34 |
0,30 |
0,38 |
|
Число пар оснований в одном витке спирали |
10,7 |
10,0 |
9,3 |
12,0 |
|
Диаметр спирали (нм) |
2,3 |
2,0 |
1,9 |
1,8 |
|
Угол наклона оснований к оси спирали (градусы) |
-1,2 |
-6 |
-9 |
|
При этом две полинуклеотидные цепочки одной молекулы являются антипараллельными, т.е. напротив 3'-конца одной цепочки находится 5'-конец другой цепочки и наоборот (рис. 4). Следует, однако, иметь в виду современные данные о том, что генетический материал некоторых вирусов представлен одноцепочечными (однонитевыми) молекулами ДНК.
На основании данных рештеноструктурного анализа
ДНК Дж. Уотсон и Ф. Крик сделали также заключение о том, что ее двухцепочечная молекула
имеет вторичную структуру в форме спирали, закрученной в направлении
слева-направо, которая в дальнейшем получила название 5-формы (рис. 5). К
настоящему времени накопились сведения о том, что помимо наиболее часто
встречающейся 5-формы можно обнаружить участки ДНК, имеющие иную конфигурацию,
- как правоза-крученную (формы А, С), так и закрученную справа-налево
(левозакрученную, или Z-форму).
2. Природа генетического кода
Необходимость кодирования структуры белков в линейной последовательности нуклеотидов мРНК и ДНК продиктована тем, что в ходе трансляции:
· нет соответствия между числом мономеров в матрице мРНК и продукте - синтезируемом белке;
· отсутствует структурное сходство между мономерами РНК и белка.
Это исключает комплементарное взаимодействие между матрицей и продуктом - принцип, по которому осуществляется построение новых молекул ДНК и РНК в ходе репликации и транскрипции.
Отсюда становится ясным, что должен существовать
"словарь", позволяющий выяснить, какая последовательность нуклеотидов
мРНК обеспечивает включение в белок аминокислот в заданной последовательности.
Этот "словарь" получил название генетического, биологического,
нуклеотидного, или аминокислотного кода. Он позволяет шифровать аминокислоты,
входящие в состав белков, с помощью определённой последовательности нуклеотидов
в ДНК и мРНК. Для него характерны определённые свойства.
.1 Триплетность
Одним из основных вопросов при выяснении свойств
кода был вопрос о числе нуклеотидов, которое должно определять включение в
белок одной аминокислоты. Сразу было понятно, что это число не может быть
равным 1 или 2, так как в этом случае количество кодирующих элементов будет
недостаточным для шифрования 20 аминокислот в белках. Число кодирующих
последовательностей из четырёх нуклеотидов по три равно 43 = 64, что более чем
в 3 раза превышает минимальное количество, которое необходимо для кодирования
20 аминокислот. В дальнейшем было установлено, что кодирующими элементами в
шифровании аминокислотной последовательности действительно являются тройки
нуклеотидов, или триплеты, которые получили название "кодоны".
.2 Смысл кодонов
Смысл кодонов стал понятен в 60-х г. XX столетия, когда, используя бесклеточную систему синтеза белков (табл. 2) и синтетические полирибонуклеотиды с заданной последовательностью нуклеотидов в качестве матрицы, М. Ниренберг и Г. Маттеи синтезировали полипептиды определённого строения. Так, на матрице поли-У, состоящей только из остатков УМФ, был получен полифенилаланин, а на матрице поли-Ц - полипролин. Из этого следовало, что триплет -UUU кодирует Фен, а триплет - ССС - Про.
В последующих экспериментах использовали смешанные синтетические полирибонуклеотиды с известным составом. В результате этой работы удалось установить, что из 64 кодонов включение аминокислот в синтезирующуюся полипептидную цепь шифрует 61 триплет, а 3 остальных - UAA, UAG, UGA не кодируют включение в белок аминокислот и первоначально были названы бессмысленными, или нон-сенс-кодонами. Однако в дальнейшем было показано, что эти триплеты сигнализируют о завершении трансляции, и поэтому их стали называть терминирующими, или стоп-кодонами.
Кодоны мРНК и триплеты нуклеотидов в кодирующей
нити ДНК с направлением от 5' к 3'-концу имеют одинаковую последовательность
азотистых оснований, за исключением того, что в ДНК вместо урацила (U),
характерного для мРНК, стоит тимин (Т).
2.3 Специфичность
Каждому кодону соответствует только одна
определённая аминокислота. В этом смысле генетический код строго однозначен.
Таблица 2. Основные компоненты белоксинтезирующей системы
|
Необходимые компоненты |
Функции |
|
1 . Аминокислоты |
Субстраты для синтеза белков |
|
2. тРНК |
тРНК выполняют функцию адаптеров. Они акцепторным концом взаимодействуют с аминокислотами, а антикодоном - с кодоном мРНК. |
|
3. Аминоацил-тРНК синтетазы |
Каждая аа-тРНК-синтетаза катализирует реакцию специфического связывания одной из 20 аминокислот с соответствующей тРНК |
|
4.мРНК |
Матрица содержит линейную последовательность кодонов, определяющих первичную структуру белков |
|
5. Рибосомы |
Рибонуклеопротеиновые субклеточные структуры, являющиеся местом синтеза белков |
|
6. АТФ, ГТФ |
Источники энергии |
|
7. Белковые факторы инициации, элонгации, терминации |
Специфические внерибосомные белки, необходимые для процесса трансляции (12 факторов инициации: elF; 2 фактора элонгации: eEFl, eEF2, и факторы терминации: eRF) |
|
8. Ионы магния |
Кофактор, стабилизирующий структуру рибосом |
Примечания: elF (eukaryotic initiation factors)
- факторы инициации; eEF (eukaryotic elongation factors) - факторы элонгации;
eRF (eukaryotic releasing factors) - факторы терминации.
.4 Вырожденность
В мРНК и ДНК имеет смысл 61 триплет, каждый из которых кодирует включение в белок одной из 20 аминокислот. Из этого следует, что в информационных молекулах включение в белок одной и той же аминокислоты определяют несколько кодонов. Это свойство биологического кода получило название вырожденности.
У человека одним кодоном зашифрованы только 2 аминокислоты - Мет и Три, тогда как Лей, Сер и Apr - шестью кодонами, а Ала, Вал, Гли, Про, Тре - четырьмя кодонами (табл. 3).
Избыточность кодирующих последовательностей -
ценнейшее свойство кода, так как она повышает устойчивость информационного
потока к неблагоприятным воздействиям внешней и внутренней среды. При
определении природы аминокислоты, которая должна быть включена в белок, третий
нуклеотид в кодоне не имеет столь важного значения, как первые два. Как видно
из табл. 4-4, для многих аминокислот замена нуклеотида в третьей позиции кодона
не сказывается на его смысле.
.5 Линейность записи информации
В ходе трансляции кодоны мРНК "читаются" с фиксированной стартовой точки последовательно и не перекрываются. В записи информации отсутствуют сигналы, указывающие на конец одного кодона и начало следующего.
Кодон AUG является инициирующим и прочитывается
как в начале, так и в других участках мРНК как Мет. Следующие за ним триплеты
читаются последовательно без каких-либо пропусков вплоть до стоп-кодона, на
котором синтез полипептидной цепи завершается.
.6 Универсальность
До недавнего времени считалось, что код
абсолютно универсален, т.е. смысл кодовых слов одинаков для всех изученных
организмов: вирусов, бактерий, растений, земноводных, млекопитающих, включая
человека. Однако позднее стало известно одно исключение, оказалось, что
митохондриальная мРНК содержит 4 триплета, имеющих другое значение, чем в мРНК
ядерного происхождения. Так, в мРНК митохондрий триплет UGA кодирует Три, AUA -
Мет, а АСА и AGG прочитываются как дополнительные стоп-кодоны.
.7 Колинеарность гена и продукта
У прокариотов обнаружено линейное соответствие
последовательности кодонов гена и последовательности аминокислот в белковом
продукте, или, как говорят, существует колинеарность гена и продукта.
Таблица 3. Генетический код
|
Первое основание |
Второе основание |
|||
|
|
U |
С |
А |
G |
|
U |
UUU Фен |
UCU Cep |
UAU Тир |
UGU Цис |
|
|
UUС Фен |
UCC Сер |
иАСТир |
UGC Цис |
|
|
UUА Лей |
UCA Cep |
UAA* |
|
|
|
UUG Лей |
UCG Сер |
UAG* |
UGG Apr |
|
С |
CUU Лей |
CCU Про |
CAU Гис |
CGU Apr |
|
|
CUC Лей |
ССС Про |
САС Гис |
CGC Apr |
|
|
CUA Лей |
ССА Про |
САА Глн |
CGA Apr |
|
|
CUG Лей |
CCG Про |
CAG Глн |
CGG Apr |
|
А |
AUU Иле |
ACU Tpe |
AAU Асн |
AGU Сер |
|
|
AUC Иле |
АСС Тре |
ААС Асн |
AGG Сер |
|
|
AUA Мет |
АСА Тре |
ААА Лиз |
AGA Apr |
|
|
AUG Мет |
ACG Тре |
AAG Лиз |
AGG Apr |
|
G |
GUU Ban |
GCU Ала |
GAU Асп |
GGU Гли |
|
|
GUC Вал |
GCC Ала |
GAC Асп |
GGC Гли |
|
|
GUА Вал |
GСА Ала |
GАА Глу |
GGA Гли |
|
|
GUG Вал |
GСG Ала |
GAG Глу |
GGG Гли |