Sl3
В последние два десятилетия найдено много образцов анти-Kn-подобных антител, в том числе указывающих на гетерогенность антигенов Sl a и Vil.
Обнаруженный с помощью одной из сывороток антиген Sl3 включен в систему Knops под обозначением KN8. Антитела анти-Sl3 выявлены у европейки, которая пока остается единственным известным индивидом, имеющим фенотип Sl(a + )Vil−Sl3 −. Других носителей анти-Sl3-антител не обнаружено.
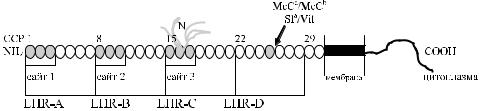
Moulds и соавт. [49] пришли к заключению, что серологические различия антигенов Sl a,Vil и Sl3 обусловлены следующими молекулярными замещениями:
позицияArg 1601 → антиген Sl 1 (Sla) позиция Gly 1601 → антиген Sl 2 (Vil) позицииArg 1601 и Ser 1610 → антиген Sl 3 позиция Ser 1610 → антиген Sl 4
позиция Tre 1610 → антиген Sl 5
Последние две позиции, как полагают указанные авторы, соответствуют гипотетическим антигенам, существование которых представляется вполне реальным.
Yka
Антитела анти-Yk a (York) сначала были приняты за анти-Cs a, поскольку реагировали с эритроцитами Cs(a + ), но не реагировали с двумя образцами эритроцитов Cs(a −). Однако миссис York, у которой впервые были выявлены антитела, была Cs(a + ) и, соответственно, не могла вырабатывать анти- Cs a-антитела. В связи с этим фактор Yk a (York) был квалифицирован как новый антиген, фенотипически ассоциированный с антигеном Cs a коллекции
Cost (Molthan, Giles [37]).
Посемейные исследования показали кодоминантное наследование антигена Yk a. Его частота составила 92 % среди европеоидов, 98 % среди негроидов
(Molthan, Giles [37], Molthan [35]).
Частота фенотипа Cs(a −)Yk(a −) среди европеоидов – 1,9 % (Molthan, Giles [37],). Если бы между антигенами Cs a и Yk a не существовало неравновесного сцепления, то указанный фенотип должен был встречаться с частотой 0,32 %, т. е. почти в 6 раз реже. Среди негроидов лица Cs(a −)Yk(a −) встречались с частотой 0,6 % – в 25 раз чаще по сравнению с расчетной величиной (0,024 %), что также свидетельствовало о сцеплении антигеновYk a и Cs a.
Molthan и Moulds [38], сообщив об открытии антигена McC a, указали, что 37 % белых американцев имеют фенотип McC(a −)Yk(a −), а 29 % – фенотип
McC(a −)Yk(a −)Cs(a −). Среди негров фенотип McC(a −)Yk(a −) составлял 2,2 %,
фенотип McC(a −)Yk(a −)Cs(a −) ‒ 17 %. Эти показатели существенно отличались от расчетных, соответствующих положению, что гены McC a, Yk a и Cs a независимы друг от друга. Фактическое число доноров с фенотипом McC(a −) в сочетании сYk(a −), Kn(a −) и Cs(a −) оказалось в 100 раз больше ожидаемого.