Материал: Биоорганика вопросы
Олиго- и полинуклеотиды: структура, физические и химические свойства
Олигонуклеотиды (до 50 звеньев) и полинуклеотиды. Связаны фосфодиэфирными связями. Используются в качестве зондов (ДНК-зонд, меченый фрагмент для гибридизации с другим участком молекулы) или праймеров.
Читают с гидроксила рибозного кольца, который свободен (не образует фосфодиэфирной связи со следующим нуклеотидом).
Вторичная структура нуклеиновых кислот: рентгеноструктурные исследования днк, положения Чаргаффа, двойная спираль и ее биологическое значение, комплементарность и взаимная ориентация цепей
Мономеры — дезоксирибонуклеозидфосфаты. Нуклеозиды: аденозин, тимидин, гуанозин, цитидин.
П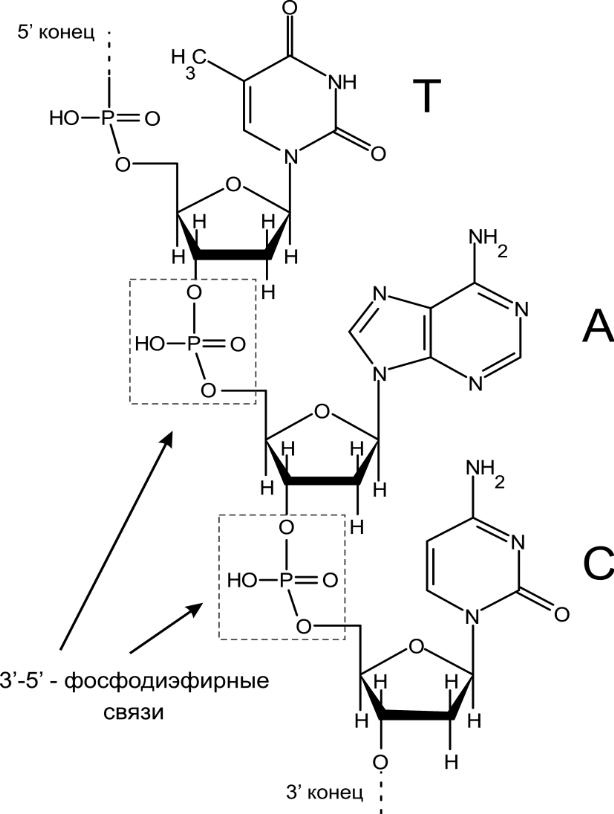 ервые
рентгеноструктурные исследования ДНК:
19 век швед Мишер обнаружил странную
субстанцию, назвал нуклеином. Уилкинс,
Франклин — четкие ренгтгеновские
дифракционные фото ДНК. Уотсон и Крик
расшифровали в 1953.
ервые
рентгеноструктурные исследования ДНК:
19 век швед Мишер обнаружил странную
субстанцию, назвал нуклеином. Уилкинс,
Франклин — четкие ренгтгеновские
дифракционные фото ДНК. Уотсон и Крик
расшифровали в 1953.
О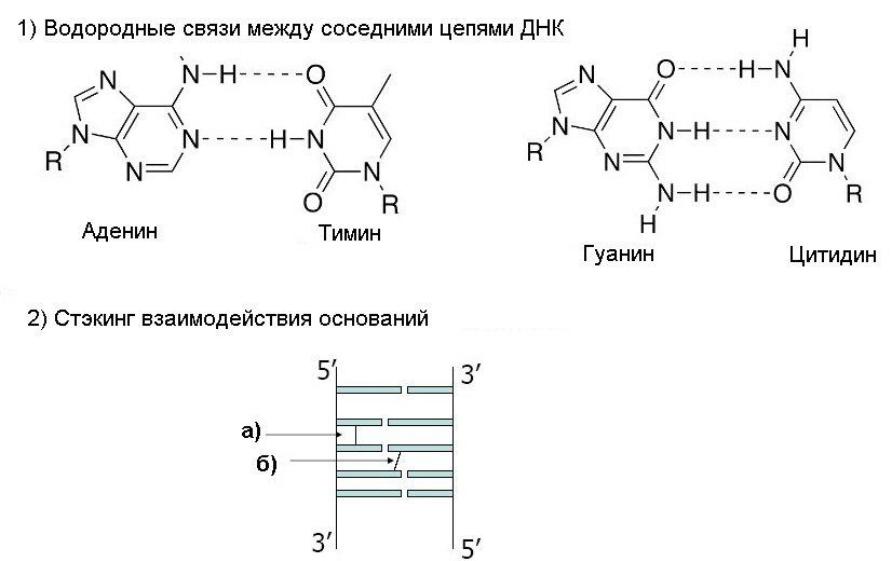 риентация
цепей:
ДНК — двойная правозакрученная спираль.
Цепи соединены водородными связями,
антипараллельны (3'-5' VS 5'-3').
риентация
цепей:
ДНК — двойная правозакрученная спираль.
Цепи соединены водородными связями,
антипараллельны (3'-5' VS 5'-3').
Комплементарность: водородные связи только между G-C (3 связи) и A-T (2 связи). В одном витке 10 нуклеотидов, высота 3,4 нм. Помимо водородных связей двойная спираль стабилизируется стэкинг-взаимодействиями между гетероциклами, которые расположены "стопками" высотой 3,4 А. До конца не ясно, что и как, но характер гидрофобный, а между облаками π-электронов ароматических колец оснований есть частичное перекрывание. Энергия 12÷60 кДж/моль. Возможно благодаря этому есть гиперхромный эффект: поглощение УФ-света отдельными нуклеотидами существенно выше, чем полинуклеотидами, а поглощение одиночных цепей выше комплементарных.
Сахарофосфатный каркас ДНК обращен к растворителю. Неэфтерифицированные остатки фосфорной кислоты имеют заряд -1 и для компенсированная электростатического отталкивания стабилизированы противоионами Na+, K+, Mg2+. Молекулы ДНК, особенно в области сахарофосфатного участка, сильно гидратированы (насыщены водой), сеть водородных связей важна для стабилизации молекулы.
В целом ДНК — длинная молекула, внутри гидрофобные остатки гетероциклических оснований (малый желобок), на поверхности гидрофильный и заряженный сахарофосфатный остов (большой желобок).
Третичная структура: молекулы ДНК в хромосомы. Упаковка с помощью белков-гистонов — "катушек", на которые наматываются нити ДНК. Гистоны богаты остатками лизина и аргинина, положительно заряжены, образуют ионные связи с отрицательно заряженными молекулами ДНК. Каждая единица катушки = 8 гистоновых белков, вокруг них ДНК совершает 1,75 оборота (146 пар нуклеотидов) → нуклеосома. Участок ДНК между нуклеосомами — линкерный, он связан с еще одним гистоном, в среднем 60 пар. Общая масса гистонов в ядре примерно равна массе ДНК, которую они упаковывают. Нуклеосомы плотно упакованы другими белками, образуют хроматин — форма существования ДНК в интерфазных клетках. Если аминогруппы боковых радикалов лизина, аргинина модифицируются ферментами и образуют производные, заряд теряется, конформация гистонов меняется, ДНК "слетает с катушки", появляются матрицы для транскрипции. Хроматин и пакует ДНК и позволяет быстро реагировать на внутри- и внеклеточные сигналы, поддерживать состояние активности и неактивности генов. Нуклеосома — нуклеопротеид.
Денатурация — плавление (разрушение водородных и других связей) под действием температуры или pH. Так как неспаренный участки ДНК поглощают свет сильнее, чем спаренные, при плавлении двойной спирали оптическая плотность раствора возрастет. Связи G-C рвать сложнее, чем A-G. Температура плавления — кол-ва расплавленной и нерасплавленной ДНК равны. При охлаждении раствора реассоциация — отжиг.
Правила Чаргаффа: А=Т, Г=Ц; пурины = пиримидины (А+Г=Т+Ц); кол-во оснований с аминогруппами в положении 6 = кол-ву оснований с кетогруппами в положении 6: А+Ц=Т+Г.
Значение спирали: двойная спираль по принципу комплементарности → почти безошибочное копирование; "почти" = возможность возникновения наследственной изменчивости → эволюции; передача наследственной инфы и бла-бла-бла.
Рнк: строение, классификация, функции
Полимеры с рибонуклеозидами: аденонин, гуанозин, цитидин, уридин. 3'-5' связи. В отличие от ДНК не только в ядре, но и в цитозоле, рибосомах. Одноцепочечные, но есть дуплексы (двухцепочечные участки), образующие шпильки (комплементарность); в шпильках РНК антипараллельны.
2 класса: кодирующая белки мРНК и не кодирующие белки РНК (тРНК и рРНК), обслуживающие процесс синтеза белка, + митохондриальная РНК (свой генетический и белково-синтетический аппарат)
мРНК: синтезируется в ядре, последовательность (матрица) для синтеза белка, закодированного в гене. На 3'-конце 100-200 аденозинфосфатных остатов (поли(А)-фрагмент), на 5'-конце модифицированный нуклеотид 7-метилгуанозин-5'-трифосфат (кэп). Через несколько десятков нуклеотидов от кэпа инициирующий кодон — триплет AUG (Met). За кодирующим участком терминирующий или стоп-кодон UGA, UUA, UAG.
т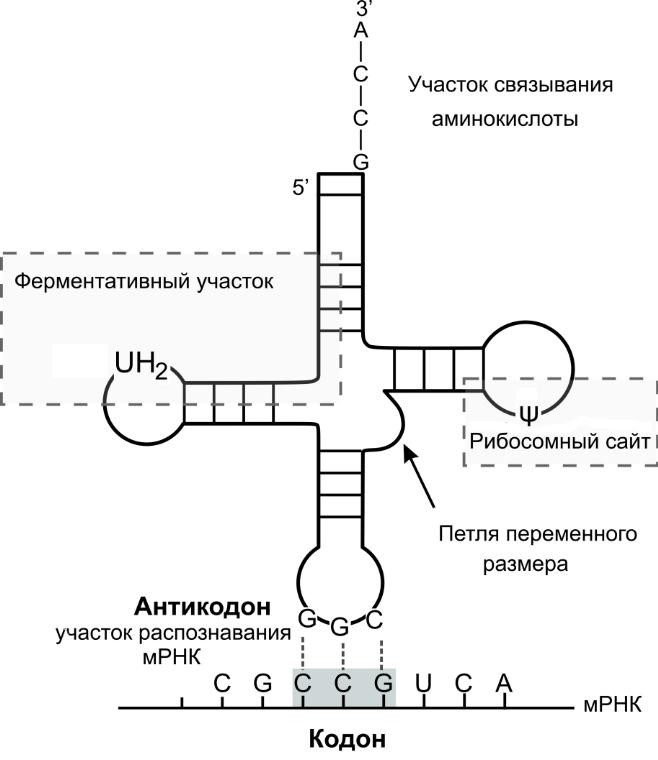 РНК:
доставка АК к рибосоме при синтезе
белка. "Клеверный лист". На 3'-конце
участок C-C-A связывания АК, на противоположном
— антикодон-триплет, комплементарный
корону матричной РНК. Каждой АК
соответствуют тРНК с триплетом-кодоном,
благодаря чему информация реализуется
однозначно. В состав тРНК входят минорные
основания (10-12 на молекулу) — метилированные
основания, изомеры, аналоги пиримидинов;
делают молекулу устойчивой к расщеплению
нуклеазами и обеспечивают пространственную
структуру, так как комплементарных пар
не образуют и препятствуют спирализации
молекулы на участках.
РНК:
доставка АК к рибосоме при синтезе
белка. "Клеверный лист". На 3'-конце
участок C-C-A связывания АК, на противоположном
— антикодон-триплет, комплементарный
корону матричной РНК. Каждой АК
соответствуют тРНК с триплетом-кодоном,
благодаря чему информация реализуется
однозначно. В состав тРНК входят минорные
основания (10-12 на молекулу) — метилированные
основания, изомеры, аналоги пиримидинов;
делают молекулу устойчивой к расщеплению
нуклеазами и обеспечивают пространственную
структуру, так как комплементарных пар
не образуют и препятствуют спирализации
молекулы на участках.
рРНК: в рибосомах, до 60% массы. Рибосомы обеспечивают трансляцию. У рРНК много удвоенных спирализованных участков, встречаются минорные основания и производные рибозы.
+ малые интерференционные РНК (миРНК, siRNA) и другие короткоцепочечные РНК, циркулирующие в крови: регуляция процессов репликации ДНК, транскрипции, трансляции.
Полимеразная цепная реакция
ПЦР — метод амплификации (накопления) нужных небольших фрагментов ДНК. Нужно для идентификации, нахождения ДНК вирусов или бактерий.
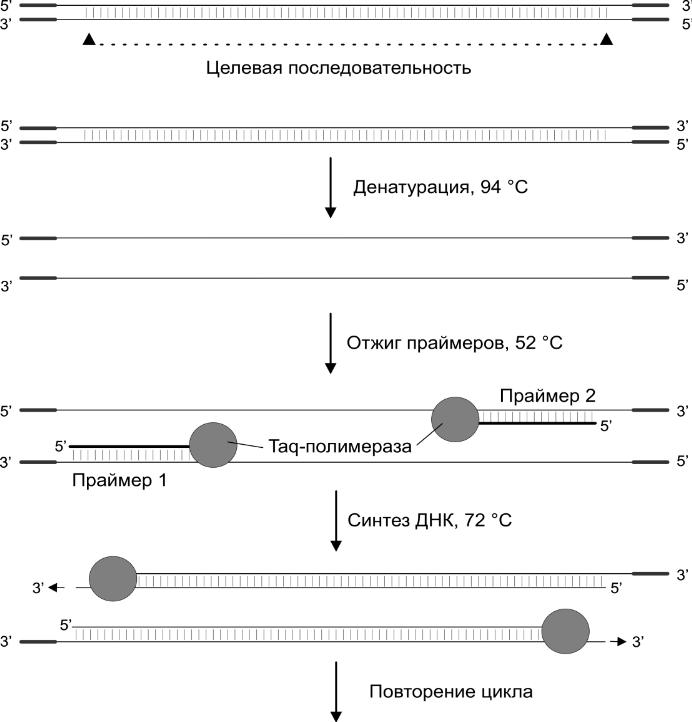
Исходные компоненты: ДНК-матрица, праймеры, дезоксинуклеотидтрифосфаты, ДНК-полимераза. Контроль с помощью температуры.
ДНК-полимераза 1 двигается по одноцепочечной молекуле ДНК, катализирует образование фосфодиэфирных связей между 3' гидроксилом предыдущего нуклеозида и 5' фосфатной группой следующего, то есть рост новой цепи от 5' к 3'. В качестве стройматериала 5' нуклеозидтрифосфаты. Скорость велика, у прокариот 1000 оснований в секунду. Полученные цепи тоже распадаются и от них цикл повторяется.
1 этап: 95, денатурация ДНК.
2 этап: 50-65, отжиг праймеров к ДНК.
3 этап: 70-72, оптимально для полимеразы — достраивание цепей праймера (Taq-полимераза).
Открыл американец Артур Корнберг.
Определение первичной структуры нуклеиновых кислот. Методы химического (по Максаму-Гилберту) и ферментативного (по Сэнгеру) с еквенирования. Автоматизация секвенирования.
Секвенирование — определение последовательности.
ХИМИЧЕСКИЙ: прочтение коротких одноцепочечных олигонуклеотидов. Гидролиз олигонуклеотида и анализ фрагментов с помощью электрофореза.
Мечение олигонуклеотида введением радиоактивного остатка фосфорной кислоты по 5'-концу. Образец делят на 4 порции и в каждую пробирку разные реагенты.
Диметилсульфат расщепляет по гуаниновым звеньям — 4 фрагмента, но только 2 радиоактивно меченые (с радиоактивным фосфатом на 5'-конце)
Муравьиная кислота по пуриновым основаниям на 8 фрагментов, 4 радиоактивные.
Гидразин по пиримидиновым, радиоактивные 4.
Гидразин в присутствии NaCl только по цитидинам — 4 фрагмента, 2 радиоактивных.
4 смеси наносят в лунки геля и проводят электрофорез, радиоактивные метки проявляют на фотобумаге.
Сборка молекулы с самого короткого фрагмента (самый большой пробег на картинке фореза).
ФЕРМЕНТАТИВНЫЙ: способность полимеразы синтезировать новую цепь нуклеотида, комплементарную материнской.
В пробирку: молекулу ДНК (матрица); 2 праймера — олигонуклеотиды длиной 20-25 оснований, комплементарные крайним участкам цепей ДНК, которые будут размножены; набор 5'-нуклеозидтрифосфатов; фермент Pol1. Праймеры нужны, потому что полимераза не работает с "пустого" места, она катализирует конденсацию между 3' гидроксилом предыдущего нуклеотида.
Денатурация ДНК
Понижение температуры, присоединение праймеров с 3' каждой цепи ("отжиг праймеров")
ДНК-полимераза синтезирует на каждой материнской цепи комплементарную дочернюю, удлиняя праймеры 5' → 3'. Полученные 2 двойные молекулы плавят, обжигают праймеры, затем они "прирастают" уже на 4 копиях. С каждым циклом в смеси кол-во копий увеличивается в 2n раз.
Полимераза в ПЦР должна сохранять активность при высокой температуре длительное время, что для белков не характерно — спасибо термофильным бактериям с Taq-полимеразой, которая активна около 100 градусов.
С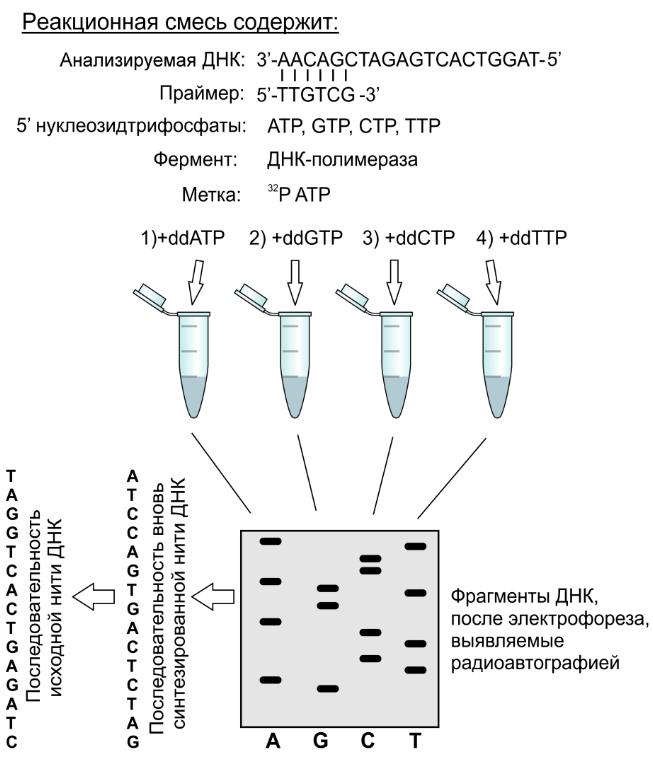 енгер
разработал способ секвенирования ДНК
на основе ПЦР: метод обрыва цепи или
метод дидезокситерминаторов.
Обрыв/терминация цепи происходит, когда
в дочернюю цепь включается нуклеозидтрифосфат
без 3'-гидроксильной группы —
дидезоксирибонуклеозидтрифосфат.
енгер
разработал способ секвенирования ДНК
на основе ПЦР: метод обрыва цепи или
метод дидезокситерминаторов.
Обрыв/терминация цепи происходит, когда
в дочернюю цепь включается нуклеозидтрифосфат
без 3'-гидроксильной группы —
дидезоксирибонуклеозидтрифосфат.
Все необходимые компоненты ПЦР
Делят на 4 части и дополнительно вносят везде ДДНКТФ
Каждый раз, когда при синтезе цепи в цепь включается остаток, синтез останавливается (нет гидроксила), поэтому в пробирке накапливаются олигонуклеотиды разной длины и заканчивающиеся либо на аденозине, либо на цитидине, либо на гуанозине, либо на тимидине.
Смеси анализируют с помощью гель-электрофореза в денатурирующих условиях. В зависимости от размера продукты ПЦР двигаются в геле с разной скоростью. Картинку проявляют радиоавтографией, для чего продукты метят изотопной меткой. Полученное изображение читают снизу-вверх по полоскам.
Итоговая последовательность комплементарна исходной матрице.
Автоматизация секвенирования: уже в автоматическом режиме, для анализа флуоресцентные метки, которые присоединяют к молекулам дидезокситрифосфатов. Каждая метка имеет свой спектр свечения. Для электрофореза используют капилляры с гелем. Свечение инициируется лазерным лучом и по цвету соотносится с основанием. Есть генный анализатор до 1000 нуклеотидов определяет, 16 образцов одновременно.
Химический синтез олигонуклеотидов: основные принципы, синтез на полимерном носителе
Сегодня с помощью автоматических синтезаторов.
Для направленного синтеза нужно блокировать функциональные группы нуклеозида, не участвующие в данном этапе синтеза. Для этого используют ацильные остатки, которые потом удаляются в щелочных условиях.
Во всех синтезаторах синтез твердофазным методом: первый нуклеозид иммобилизован на поверхности нерастворимых полимерных частиц, а растущая цепочка экспонирована к реакционной смеси с реагентами. Частицы упакованы в колоночных реакторах, через которые последовательно прокачиваются растворы веществ для синтеза. Для оптимальных гидродинамических параметров колонки частицы имеют сферическую форму и одинаковые размеры. Ключевые мономеры для синтеза часто 4 производные нуклеозид фосфорамидита, где все ФГ заблокированы защитными. Конденсация проходит в течение нескольких минут. Экзоцикличные аминогруппы в составе гетероциклических оснований заблокированы группировками (ацетильными остатками), которые потом удаляются в щелочных условиях. Такие мономеры — синтоны (реальные или идеализированные структурные единицы молекул, которые можно ввести в химических синтез).
Общая схема:
О
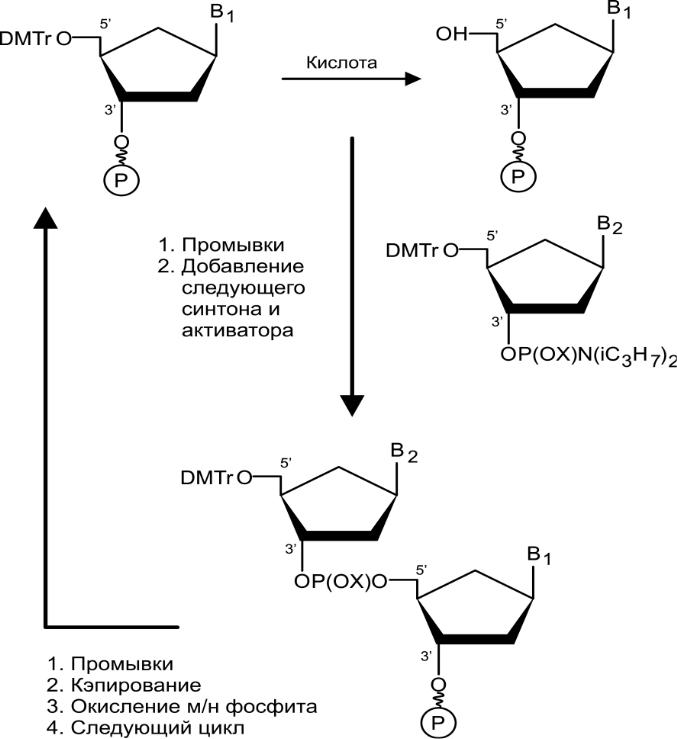 свобождение
кислотной обработкой 5'-гидроксила
первого нуклеозида (иммобилизован)
и промывка реактора (детрилирование).
свобождение
кислотной обработкой 5'-гидроксила
первого нуклеозида (иммобилизован)
и промывка реактора (детрилирование).Внесение в реактор смеси следующего мономера — синтона и активатора (тетразола), присоединение фосфорамидита (удлинение цепи на звено).
Промывки, введение в реактор уксусного ангидрида для цацетилирования непрореагировавших гидроксильных групп (кэпирования).
Промывки, окисление межнуклеотидного фосфита до фосфата.
И так по циклам, каждый 5-7 минут. Для высокого выхода он должен быть высоким на каждом цикле. Повышению эффективности способствует высокая степень чистоты реагентов и химикатов, поэтому важно кэипирование, блокирование гидроксильных групп, чтобы не накапливались ложные последовательности. После синтеза ЗГ удаляют, полученный продукт отсоединяют от частицы.
БИОХИМИЯ БИЛЕТЫ №13-17 (углеводы)
М
 оносахариды:
определение, номенклатура, линейные и
циклические формы моносахаридов,
стереохимия и конформация моносахаридов.
Аномерный центр: его стереохимия, особые
свойства гидроксильной группы.
оносахариды:
определение, номенклатура, линейные и
циклические формы моносахаридов,
стереохимия и конформация моносахаридов.
Аномерный центр: его стереохимия, особые
свойства гидроксильной группы.
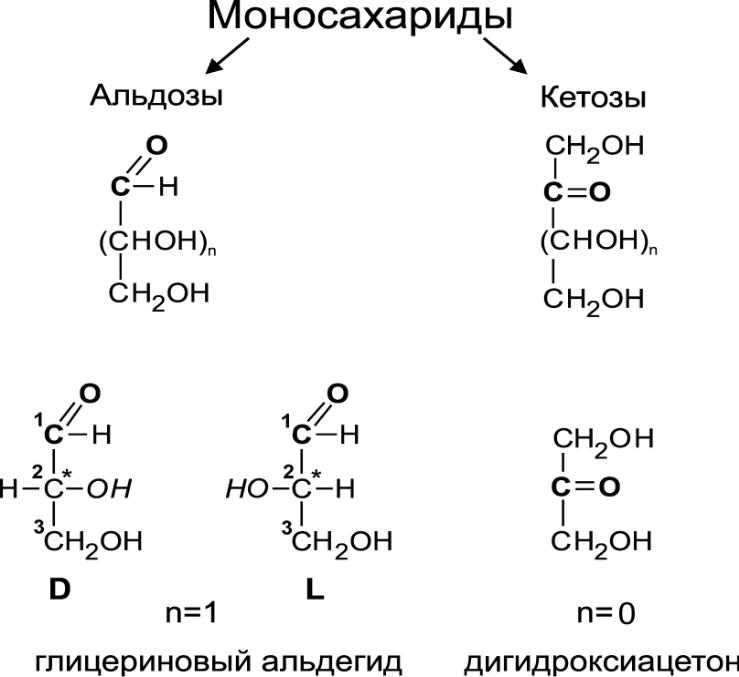
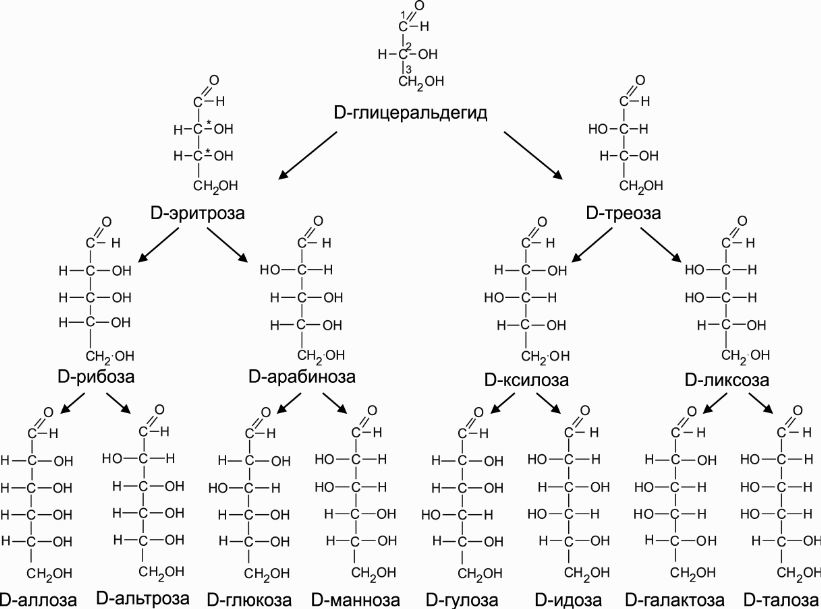
Cn(H2O)n, n ≥ 3.
Моносахариды — молекулы, в составе которых одна или несколько гидроксильных групп и карбонильная группа. Если карбонил альдегидный, тогда альдоза, если кетонный, то кетоза. По кол-ву С-атомов: треоза, тетроза, пентоза...
С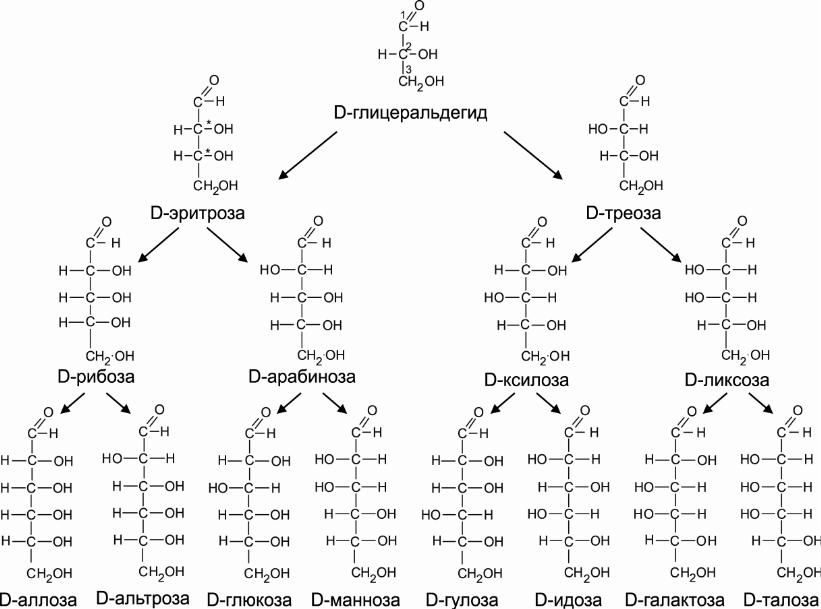
 амые
маленькие: глицериновый альдегид =
альдотриоза; дигидроксиацетон.
амые
маленькие: глицериновый альдегид =
альдотриоза; дигидроксиацетон.
К ол-во стереоизомеров: 2n (n — кол-во хиральных центров). Хиральный центр — H-C-OH (все заместители разные). Гидроксил слева — L, гидроксил справа — D. Большинство природных углеводов с D-конфигурацией. Принадлежность определяют по гидроксилу, максимально удаленному от карбонильной группы.