Материал: Билеты по физиологии+формулы+показатели организма
36 При раздр нервной ткани был зарегистрирован пд, амплитуда к-рого по мере увеличения силы разд-я увеличивалась, а характер кривой не менялся.
Объектом был смешанный нерв.
Если электрод расположен так, что контактирует с целым нервом, то при стимуляции нерва на некотором расстоянии от места отведения электрод сначала зарегистрирует потенциалы действия наиболее быстро проводящих волокон, а после этого–группы потенциалов других, более медленно проводящих волокон. Следовательно, потенциал действия такого нерва состоит из ответов целого спектра групп волокон с разными скоростями проведения.
37 При надпороговом раздр-ии двиг нерва были зарегистрированы его биопотенциалы ( ПД) и ПД иннерв-й скелетной мышцы. ПД мышцы не возникли при действии ионов Mg, явл антагонистами ионов Са
Возбуждение передается посредством синапса, его основные составляющие – пресинаптическая мембрана, синаптическая щель, постсинаптическая мембрана
Mg не действует на первое звено передачи, так как не возник ПД на мышце. Значит Мg не действует на пресинаптическую мембрану нервного окончания.
Не действует ни на одно звено
Mg не оказывает действия на синапс, а Са действует на пресинаптическую мембрану
Са играет ключевую роль в высвобождении квантов медиатора в синаптическую щель-
Деполяризация пресинаптической мамбраны – открытие Са-каналов – высвобождение медиатора
Высвобождается медиатор, который связывается с лиганд-зависимыми Na-каналами на постсинаптической мембране, открывая их – увеличение проницаемости для натрия – генерация пд на постсинаптической мембране
38 . При надпороговом раздр двиг нерва были зарег его биопотенциалы ( ПД) и ПД иннерв-й скелет мышцы. ПД мышцы не возн при действии магния, явл антагонистом Са. При прямом раздр мышцы она продолжала возбуждаться и сокращаться
Да
Ионы магния не принимают участия в генерации пд
Нет
Так как мышца продолжала возбуждаться и сокращаться при ее прямом радражении
39 В экспер регистрировали биопотенциалы нерв вол при его пороговом раздражении и потенц иннервируемого мыш волокна при прямом раздражении. Сила раздр в обоих случаях одинакова
40 В эксперименте на 2х мышцах ( а и б ) отдельное мыш волокно раздражали внутриклеточно надпороговым током. Было установлено, что на мышце а – регистр пд и других мыш волокон, на мышце б – пд только раздражаемого волокна
Между клетками волокон существует система межклеточных контактов(нексусы), через которые распространяется пд
А – гладкая висцеральная мышца(унитарная), Б – скелетная
В гладкой мышце нет концевых пластиной и отдельных нервных окончаний, по всей длине разветвлений адрен- и холинэргических нейронов имеются варикозные утолщения, содержацие гранулы медиатора. Поэтому по ходу следования нервного волокна могут возбуждаться многие гладкомышечные клетки, а клетки не имеющие контакта с варикозами возбуждаются посредством пд, распространяющегося на них через нексусы.
В скелетной мышце нет связей между отдельными мышечными волокнами, поэтому пд не распространяется на соседние волокна
№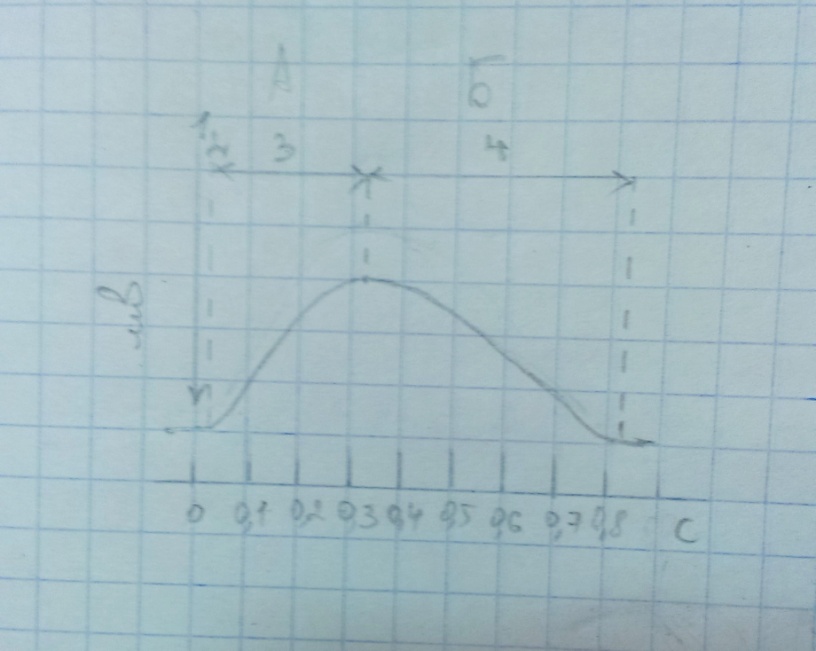 41.
Задачей эксперимента являлось получение
гладкого и зубчатого тетануса изолированной
мышцы. Длительность латентного периода
ее сокращения составляла 0.02с, периода
укорочения – 0.30с, периода расслабления
– 0.50с. Длительность фазы абсолютной
рефрактерности данной мышцы – 0.35с, фазы
относительной рефрактерности – 0.10с.
Каковы результаты эксперимента?
41.
Задачей эксперимента являлось получение
гладкого и зубчатого тетануса изолированной
мышцы. Длительность латентного периода
ее сокращения составляла 0.02с, периода
укорочения – 0.30с, периода расслабления
– 0.50с. Длительность фазы абсолютной
рефрактерности данной мышцы – 0.35с, фазы
относительной рефрактерности – 0.10с.
Каковы результаты эксперимента?
1. 1 – момент раздражения, 2 – латентный период, 3 – период укорочения, 4 – период расслабления.
2. Общее условие получения тетанического сокращения – действие на мышцу ритмических раздражений с такой частотой, что их эффекты суммируются, наступает сильное и длительное сокращение мышцы. ИЛИ наличие абсолютного рефрактерного периода (как сказано в методичке) (?)
3. При нанесении последующего раздражения во время укорочения возникает гладкий тетанус, а во время расслабления – зубчатый тетанус.
4. Минимальный промежуток времени между последовательными эффективными стимулами во время тетануса не может быть меньше рефрактерного периода. Следовательно, для данной мышцы возможно получение зубчатого тетануса (период расслабления > рефрактерный период), и невозможно получение гладкого (период укорочения < рефрактерный период).
5. Одинаковы.
6. При пороговой силе раздражителя ответ не будет наблюдаться ни во время фазы абсолютной рефрактерности, ни во время относительной. А при максимальной силе возможен будет ответ во время фазы относительной рефрактерности. Однако на частоту раздражения данной мышцы это не повлияет, так как останется: период расслабления > рефрактерный период и период укорочения < рефрактерный период.
№42. Сопоставьте динамическую работу, выполненную изолированной мышцей лягушки при однократном подъеме грузов в 10г, 30г, 410г, при условии, что абсолютная сила мышцы составляет 400г.
1. Динамическая работа – работа мышцы, при которой происходит перемещение груза и движение костей в суставах.
2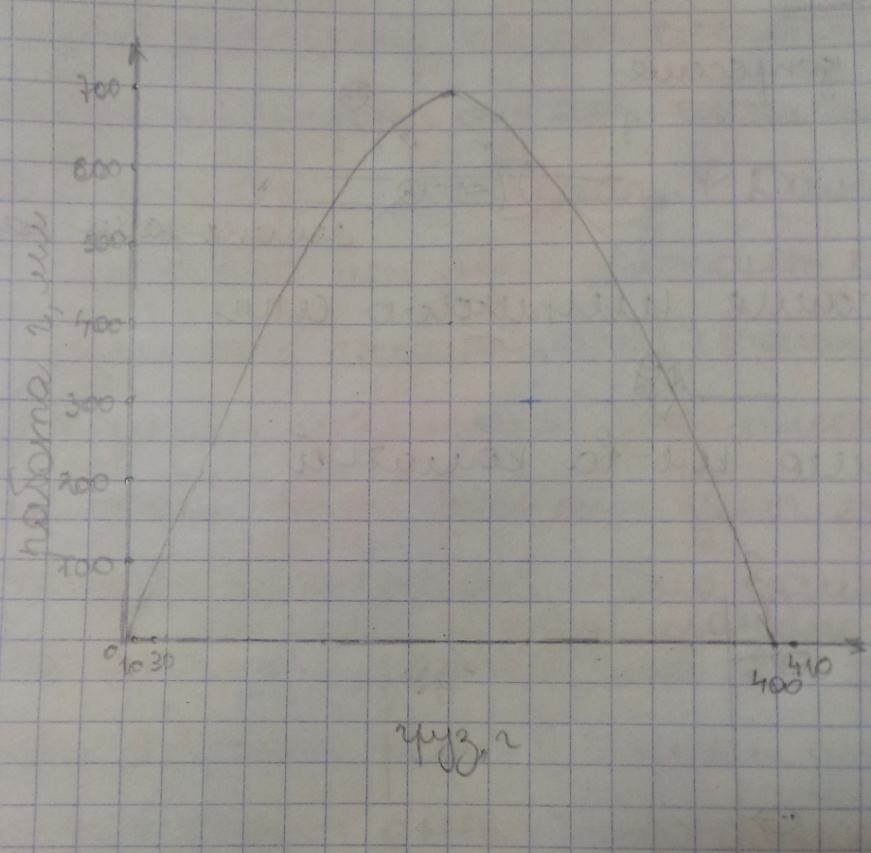 .
Учитывая, что абсолютная сила мышцы
составляет 400г, а необходимо поднять
груз 410г, то в данном случае выполненная
работа будет равняться нулю. Поэтому
при грузе 10г работа будет больше, чем
при грузе 410г.
.
Учитывая, что абсолютная сила мышцы
составляет 400г, а необходимо поднять
груз 410г, то в данном случае выполненная
работа будет равняться нулю. Поэтому
при грузе 10г работа будет больше, чем
при грузе 410г.
3. Работа при грузе 30г больше работы при грузе 10г. Высота подъема увеличилась.
4. Исходя из формулы для измерения динамической работы мышц (А = РН, где А – динамическая работа мышцы, кгм, Р – масса груза, кг, Н – высота подъема груза, м.), динамическая работа мышцы прямо пропорциональна массе груза и высоте подъема груза.
5. По мере увеличения груза работа сначала увеличивается, а затем постепенно уменьшается. При очень большом грузе, который мышца неспособна поднять, работа становится равной нулю. Наибольшую работу мышца совершает при некоторых средних нагрузках.
№43. На скелетной мышце были проведены 2 опыта (№1 и №2), в которых регистрировали ее потенциалы возбуждения (ПД). В опыте №1 на мышцах наносили прямое раздражение, в опыте №2 – непрямое. Частота раздражения в двух случаях была одинаковой и составляла 250 стимулов в секунду. Будут ли наблюдаться в опытах №1 и №2 различия в частоте ПД, возникающие в мышце при условии, что лабильность нерва равна 500 имп/с, лабильность мышцы – 300 имп/с.
1. При прямом раздражении мышцы стимулами указанной частоты, примерная частота импульсов, возникающих в мышце –250 имп/с.
2. Поскольку частота прямых раздражений - 250 стимулов в секунду, а лабильность (способность ткани воспроизводить определенное число волн возбуждения в единицу времени в точном соответствии с ритмом наносимых раздражений) мышцы – 300 имп/с.
3. При частоте раздражения нерва 250 стимулов/с, в нерве будет возникать 250 имп/с, поскольку его лабильность - 500 имп/с.
4. При непрямом раздражении указанной частоты в мышце будет возникать около 100-150 имп/с.
5. В данном случае происходит раздражение двигательного нерва, возбуждение от которого передается на мышцу посредством синапса, лабильность которого 100-150 имп/с.
6. Лабильность структур, по которым проходит возбуждение при непрямом раздражении мышцы: нерв – 500имп/с, синапс – 100-150 имп/с, мышца - 300 имп/с.
№44. Животному было введено курареподобное вещество, действующее в области мышечно-нервного синапса скелетной мышцы. Как доказать точную локализацию действия веществ и его механизм?
1. После введения примененного вещества скелетная мышца не будет реагировать на непрямое надпороговое раздражение.
2. Чтобы доказать, что указанное вещество не действует на двигательный нерв, а именно в области синапса, нужно измерить ПД отведением с данного нерва.
3. Доказать, что данное вещество не действует на сократительный аппарат мышц можно прямым раздражением.
4. Проверить, не действует ли данное вещество в области пресинаптической мембраны необходимо, проверив наличие медиатора ацетилхолина в синаптической щели.
5. Проверить, не оказывает ли данное вещество антихолинэстеразное действие, можно измерив ПД постсинаптической мембраны, т.к. при действии ингибитора холинэстеразы ритмическое раздражение нерва вызывает суммацию потенциала концевой пластинки, что ведет к стойкой деполяризации постсинаптической мембраны. (?)
6. Обладает ли вещество деполяризующим действием нужно проверить, измерив ПД постсинаптической мембраны. (?)
7. Проверить, обладает ли вещество конкурентным действием, можно искусственным введением ацетилхолина. (?)
8. На основании проведенного анализа остается получить результат: данное вещество воздействует на холинорецепторы – курареподобные вещества блокируют их, нарушая открытие Na+каналов на постсинаптической мембране.
№45. При раздражении нерва, иннервирующего мышцу, была зарегистрирована гиперполяризация постсинаптической мембраны. К какому виду относится раздражаемая мышца?
1. Описанное изменение заряда потенциала соответствует утомлению мышцы.
2. Следовательно, синапсы, возбуждаемые при раздражении данного нерва – тормозные.
3. Нервные импульсы, приходящие в варикозы аксонов, высвобождают тормозной медиатор. Воздействуя на постсинаптическую мембрану, тормозной медиатор взаимодействует с хемовозбудимыми каналами, обладающими преимущественной проницаемостью для ионов К+. Выходящий поток калия через эти каналы вызывает гиперполяризацию постсинаптической мембраны.
4. Раздражаемая мышца – гладкая, т.к. именно гладкие мышцы могут возбуждаться и тормозиться благодаря наличию адренергических и холинергических волокон.
№46. В эксперименте на нервно-мышечном препарате лягушки регистрировали биопотенциалы и сокращение мышечного волокна при увеличении частоты его раздражения от 20 до 200 стимулов в секунду. Использовали два способа раздражения мышц – прямое и непрямое. При одном способе раздражения увеличение частоты сопровождалось увеличением амплитуды сокращения и частоты биопотенциалов; при другом способе при увеличенной частоте раздражения мышца расслаблялась, биопотенциалы не регистрировались.
Объясните результат эксперимента при условии, что лабильность раздражаемой мышцы составляла 250 имп/с.
1. При непрямом раздражении мышечного волокна раздражают двигательный нерв, возбуждение по которому посредством синапса передается на мышцу.
2. Доказать, что лабильность мышцы 250 имп/с можно, увеличивая частоту ее раздражения от 200 до 300 стимулов/с. На раздражение 250 и меньше стимулов/с мышца будет отвечать соответствующим числом сокращений в единицу времени в точном соответствии с ритмом наносимых раздражений. Если частота раздражения превышает величину лабильности, то возникает ее трансформация.
3. Лабильность структур при непрямом раздражении: самая большая – нерв, меньше – мышца, еще меньше – синапс.
4. При частоте прямого раздражения 20 имп/с и 200 имп/с мышца будет генерировать ПД с соответствующей частотой. При частоте непрямого раздражения 20 имп/с мышца будет генерировать ПД соответствующей частотой, а при частоте 200 имп/с – с меньшей частотой.
5. Явление уменьшения частоты биопотенциалов и амплитуды мышечного сокращения вплоть до прекращения генерации ПД и расслабление мышц при увеличении частоты раздражения – утомление.
6. При длительном раздражении в нервных окончаниях уменьшается запас медиатора, а его синтез не поспевает за расходованием. Накапливающиеся продукты обмена в мышце понижают чувствительность постсинаптической мембраны к ацетилхолину, в результате чего уменьшается величина постсинаптического потенциала. Когда он понижается до критического уровня, в мышечном волокне не возникает возбуждения. Снижается скорость ресинтеза АТФ, необходимого для мышечного сокращения.
№47. Задачей эксперимента на двух мышцах (А и Б) являлось получение суммарного мышечного сокращения. Предварительно было установлено, что длительность периода укорочения мышцы А – 50 мс, мышцы Б – 500 мс.
1. Для получения слитного суммарного сокращения мышцы А и Б необходимо их раздражать с высокой частотой.
2. Для возникновения суммации необходимо, чтобы интервал между раздражением имел определенную длительность: он должен быть длиннее рефрактерного периода, иначе на второе раздражение не будет ответа, и короче всей длительности сократительного ответа, чтобы второе раздражение подействовало на мышцу раньше, чем она успеет расслабиться после первого раздражения. Причем для возникновения слитного суммированного сокращения раздражение должно приходиться в фазу укорочения.
3. Слитное сокращение мышцы А и мышцы Б – гладкий тетанус.
4. Мышца А относится к медленным красным, Б – к быстрым белым.
5. Продолжительность суммарного сокращение мышцы А будет большей, нежели мышцы Б, т.к. медленные мышечные волокна обладают большей выносливостью, чем быстрые.
№48. На скелетную мышцу лягушки наносили непрямое раздражение и регистрировали сокращения при частоте 5, 15, 25, 50 и 150 стимулов в секунду. Каковы результаты раздражения?
1. Ответ мышцы на 5 стимулов в секунду – 5 одиночных сокращений, поскольку при воздействии на мышцу непрямых раздражений с большим интервалом, при котором всякое следующее раздражение попадает в период после окончания фазы расслабления.
2. Ответ мышцы на 15 стимулов в секунду – зубчатый тетанус, так как подается много стимулов с интервалом, при котором после первого раздражения сокращение еще не закончилось, а уже возникло новое.
3. Ответ мышцы на 25 стимулов в секунду – зубчатый тетанус, поскольку подается много стимулов с интервалом, при котором после первого раздражения сокращение еще не закончилось, а уже возникло новое.
4. Ответ мышцы на 50 стимулов в секунду – гладкий тетанус, т.к. стимулы наносятся с очень малым интервалом. При сравнении с раздражением 25 стимулов/с последующее раздражение приходится на фазу укорочения, а не расслабления.
5. На непрямое раздражение в 150 имп/с мышца ответит гладким тетанусом, т.к. стимулы наносятся с очень высокой частотой.
6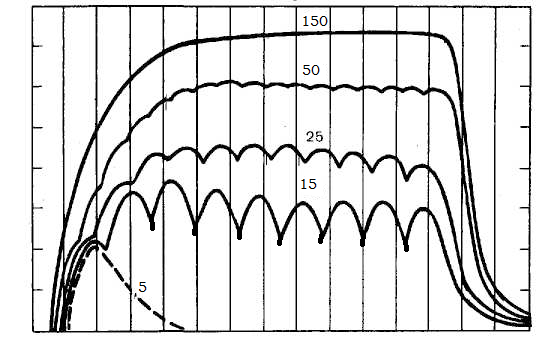 .
.
№49. Икроножную мышцу и мышцу желудка лягушки помещали в раствор, содержащий ионы Na+, K+, Ca2+ и Cl- в концентрациях, соответствующих их содержанию в тканях. На мышцы наносили прямое надпороговое раздражение и регистрировали биопотенциалы и сокращения.
Как отразилось на результатах эксперимента прибавление к раствору ионов Mg2+, который, как известно, обладает высоким сродством к активным группам кальциевых каналов и, связываясь с ними, блокирует прохождение Ca2+?
1. При блокаде кальциевых каналов нарушается мышечное сокращение.
2. Прибавление ионов магния не повлияло на генерацию ПД икроножной мышцы.
3. Генерация ПД происходит благодаря ионам Na+, К+ и не зависит от ионов Ca2+.
4. Блокада кальциевых каналов повлияла на сокращение скелетной мышцы – его осуществление стало невозможным.
5. Ионы Ca2+ необходимы для сокращения мышцы: они присоединяются к тропонину, и тот прекращает блокировать взаимодействие актина с миозином.
6. Прибавление ионов магния повлияло на генерацию ПД мышцей желудка во время ее раздражения – заблокировало ее.
7. Ионная природа ПД гладкой мышцы определяется особенностями каналов мембраны гладкой мышечной клетки. Основную роль в механизме возникновения ПД играют ионы Са2+.
8. При действии ионов Mg2+ сократимость мышцы желудка изменилась – стала отсутствовать.
9. Ионы Ca2+ перестали поступать в цитоплазму миоцитов желудка. Данные ионы должны связываться с кальмодулином, активирующим киназу легкой цепи миозина, которая приводит к гидролизу АТФ и запускает цикл образования поперечных мостиков.
10. Прибавление ионов магния не влияет на генерацию ПД исчерченной мышцы, однако блокирование кальциевых каналов данными ионами нарушает способность мышцы сокращаться. В гладкой мускулатуре блокирование кальциевых каналов ионами Mg2+ негативно воздействует и на ПД, и на сокращение.
№50. Изолированные икроножные мышцы одной и той же лягушки раздражали с частотой 8 имп/с. Одна из мышц была предварительно утомлена, остальные условия проведения эксперимента были одинаковыми. Оказалось, что сокращения одной из мышц были одиночными, другая отвечала суммарными сокращениями.
Какая мышца отвечала на раздражения одиночными, какая – суммарными сокращениями?
1. Неутомленная мышца не может в данных условиях отвечать на раздражение суммарным сокращением, ведь осуществляется воздействие на мышцу раздражений с большим интервалом, при котором всякое следующее раздражение попадает в период после окончания фазы расслабления.
2. Утомленная мышца может в данных условиях отвечать на раздражение суммарным сокращением. При утомлении нарастает латентный период сокращения и удлиняется период расслабления мышцы, следовательно, последующие сокращения могут попадать в фазу расслабления, вызывая суммарное сокращение.
3. Вывод: неутомленная мышца отвечала на раздражения одиночными, а утомленная – суммарными сокращениями.
№ 1 Известно, что ионные каналы мембраны возбудимой клетки регулируют амплитуду мембранных потенциалов. Экспериментально обнаружено, что яд тетродотоксин блокирует натриевые каналы мембраны возбудимой клетки. Вопросы: 1. Как изменится при этом потенциал покоя возбудимости клетки? 2. Как изменится при этом потенциал действия возбудимой клетки? 3. Как при этом изменится распределение ионов на внешней и внутренней стороне клеточной мембраны? 4. Повлияет ли тетродотоксии на проведение возбуждения по нервному волокну?
1. Известно, что потенциал покоя создается преимущественно за счет выхода ионов калия по концентрированному градиенту из клетки. При этом натриевые каналы частично открыты, и некоторое количество ионов натрия проходит в клетку, уменьшая потенциал покоя. Следовательно, блокада натриевых каналов тетродотоксином приведет к небольшому увеличению потенциала покоя. 2. При блокаде натриевых каналов становится невозможным возникновение потенциала действия, так как деполяризация клеточной мембраны невозможна. 3. Концентрация ионов натрия на внешней стороне клеточной мембраны незначительно увеличится, так как ионы натрия полностью перестают входить в клетку. 4. Поскольку проницаемость натриевых каналов увеличивается при возникновении потенциала действия, их блокада тетродоксином приведёт к невозможности распространения возбуждения по нервному волокну. |
№ 2 Известно, что фазы потенциала действия нервного волокна (быстрая деполяризация и реполяризация) возникают вследствие движения ионов натрия и калия вдоль концентрационных градиентов. В эксперименте на нерв подействовали уабаином — веществом, подавляющим активность АТФазы, затем провели длительное ритмическое раздражение нерва. Вопросы: 1. Как при этом изменится распределение ионов на внешней и внутренней стороне клеточной мембраны? 2. Изменится ли величина потенциала покоя и потенциала действия в обработанном уабаином нервном волокне?
1. Потенциал покоя и потенциал действия возбудимых клеток обусловлены разной концентрацией ионов, в первую очередь, калия и натрия, снаружи и внутри клетки. Разность концентраций ионов поддерживается благодаря калиево-натриевому насосу, работа, которая является энергозависимой и требует АТФазной активности. Следовательно, ингибирование АТФазы приведет к выравниванию концентраций калия и натрия снаружи и внутри нервных волокон в ходе ритмического раздражения нерва. 2. Отсутствие градиента концентрации ионов снаружи и внутри клетки приведет к исчезновению потенциала покоя и полной невозможности возникновения потенциала действия. |
№ 3 Известно, что градиент концентрации натрия между двумя сторонами клеточной мембраны влияет на величину потенциалов покоя и действия возбудимой клетки. В эксперименте увеличили концентрацию ионов натрия внутри нервной клетки вначале весьма незначительно, затем выровняли концентрацию ионов натрия внутри клетки и в окружающей клетку среде. Вопрос: Как изменится величина потенциалов покоя и действия в двух описанных ситуациях?
1. Небольшое увеличение концентрации положительно заряженных ионов натрия внутри клетки уменьшит разность потенциалов снаружи и внутри клетки, что вызовет небольшую деполяризацию. Разность потенциалов приблизится к критическому уровню деполяризации, следовательно, возбудимость повысится, и потенциалы действия будут возникать легче. 2. При выравнивании концентрации ионов натрия внутри клетки и снаружи клетки потенциал действия не сможет возникнуть, поскольку натрий не сможет входить в клетку в условиях отсутствия градиента концентрации. |
|
|
|
||
|
|
||
№ 4 Известно, что препарат этилендиаминтетраамоний (ЭДТА) связывает в живых тканях ионы кальция. В экспериментальных условиях на нервно-мышечном препарате лягушки в область нервно-мышечного синапса ввели ЭДТА. Вопросы: 1. Как изменится процесс проведения возбуждения в синапсе? 2. Как повлияет ЭДТА на синтез ацетилхолина, проницаемость пресинаптической мембраны для медиатора, генерацию постсинаптического потенциала и активность холинэстеразы?
При распространении возбуждения нервного волокна в пресинаптическую область увеличивается проницаемость пресинаптической мембраны и ионы кальция входят по градиенту концентрации внутрь волокна. Связываясь с пресинаптическими везикулами, кальций обеспечивает движение везикул в сторону пресинаптической мембраны, что необходимо для высвобождения медиатора в синаптическую щель. 1. Связывание ионов кальция ЭДТА приведет к прекращению высвобождения ацетилхолина в нервно-мышечном синапсе и блокаде проведения через синапс. 2. ЭДТА не повлияет непосредственно на синтез ацетилхолина, проницаемость пресинаптической мембраны для медиатора, генерацию постсинаптического потенциала и активность холинэстеразы. |
№ 5 Известно, что проведение возбуждения в синапсе состоит из нескольких стадий. В эксперименте воздействие химического вещества на нервно-мышечные синапсы привело к прекращению передачи возбуждения с нерва на скелетную мышцу. При введении в указанную область ацетилхолина проведение возбуждения через синапс не восстановилось. Введение фермента ацетилхолинэстеразы восстановило проведение возбуждения. Вопросы: 1. Перечислите возможные механизмы прекращения проведения возбуждения в синапсе? 2. Каков механизм действия изучаемого вещества на нервно-мышечный синапс?
1. Возможными причинами прекращения перехода возбуждения с нерва на мышцу в синапсе могут быть: нарушение выделения медиатора ацетилхолина пресинаптической областью; инактивация или блокада холинэргических рецепторов постсинаптической мембраны; ингибирование фермента, разрушающего ацетилхолин. 2. При добавлении ацетилхолина нервно-мышечная передача не восстановилась, следовательно, дело не в недостаточном выделении медиатора. Добавление фермента холинэстеразы, расщепляющего ацетилхолин и освобождающего рецепторы постсинаптической мембраны для взаимодействия со следующими квантами медиатора, восстановило синаптическую передачу. Следовательно, изучаемое вещество является ингибитором холинэстеразы. |
№ 6 Известно, что гладкие мышцы имеют ряд физиологических особенностей по сравнению со свойствами скелетных мышц. В ходе эксперимента из стенки кишечника и стенки артерии мышечного типа животного было выделено по фрагменту (длиной 2 см и шириной 2 см), содержащему гладкомышечные волокна. Третий фрагмент такого же размера был выделен из скелетной мышцы. Внешне мало отличающиеся друг от друга мышечные фрагменты поместили в камеру с физиологическим раствором, что обеспечивало условия для их жизнедеятельности в течение некоторого времени. Вопросы: 1. Как различить принадлежность фрагментов мышечной ткани по их функциональным свойствам? 2. По какому функциональному признаку, без применения воздействий, можно идентифицировать принадлежность одного из фрагментов к мышечной ткани кишечника? 3. Как с помощью раздражения фрагментов мышечной ткани можно отличить мышечную ткань внутренних органов от скелетной мышцы?
1. При наличии морфологического сходства между тремя фрагментами мышечной ткани, фрагменты гладкомышечной ткани отличаются автоматизмом, т.е. способностью к спонтанной генерации потенциалов действия и к сокращению. 2. Автоматия хорошо выражена у гладких мышц стенок полых органов, в частности, кишечника, и нехарактерна для гладких мышц стенок кровеносных сосудов. 3. Гладкие мышцы в отличие от скелетных мышц обладают высокой чувствительностью к биологически активным веществам. Подведение раствора ацетилхолина вызовет ритмические сокращения мышечного фрагмента кишечника. Раствор адреналина вызовет спастическое сокращение фрагмента скелетной мышцы. Сокращение фрагмента скелетной мышцы можно вызвать электрическим раздражением. |
|
|
|
||
|
|
||
№ 7 Известно, что одним из основных свойств возбудимых тканей является возбудимость. Экспериментально сравнивали возбудимость нервной и мышечной ткани до и после длительного прямого и непрямого раздражения мышцы. Было установлено, что исходно возбудимость одной ткани выше, чем второй. Кроме того, было зафиксировано изменение возбудимости нерва и мышцы после длительного раздражения. Вопросы: 1. Как определялась возбудимость нерва и мышцы? 2. Какая ткань и почему имела большую возбудимость? 3. Как изменилась возбудимость нерва и мышцы после длительного прямого и непрямого раздражения мышцы? 4. Какие параметры характеризуют величину возбудимости ткани?
1. На нервно-мышечном препарате лягушки сначала раздражали нерв и затем мышцу одиночными электрическими импульсами. Силу раздражения постепенно увеличивали до появления первого мышечного сокращения. Таким образом, определяли порог раздражения нерва и мышцы. 2. Нервная ткань имела большую возбудимость, так как ее порог раздражения был ниже по сравнению с мышечной тканью. 3. Возбудимость понизится вследствие уменьшения соотношения ионов на клеточной мембране при длительном раздражении нерва и мышцы. 4. Величина возбудимости ткани характеризуется порогом раздражения, реобазой, хронаксией, скоростью аккомодации. |
№ 8 Известно, что возбудимость является одним из основных свойств нервной и мышечной тканей. Экспериментально было установлено, что после нанесения на изолированный нерв и полоску миокарда надпорогового раздражения возникали потенциалы действия, в ходе которых происходило изменение возбудимости. Вопросы: 1. Какой метод регистрации использовали для изучения потенциалов действия в нерве и полоске миокарда? 2. Как изучали изменение возбудимости в различные фазы потенциалов действия? 3. Как изменяется возбудимость при возникновении потенциала действия?
1. Биопотенциалы регистрировали с помощью микроэлектродной техники. 2. Изменения возбудимости изучали с помощью измерений порога раздражения в различные фазы потенциалов действия. 3. Во время предспайка возбудимость повышается. При пике потенциала действия возникает абсолютный рефрактерный период. При реполяризации возникает относительный рефрактерный период. При отрицательном следовом потенциале наблюдается фаза экзальтации. При положительном следовом потенциале возникает фаза субнормальной возбудимости. |
№ 9 Известно, что вокруг клеточных мембран возбудимых тканей существует неравномерное распределение ионов. Экспериментально увеличивали градиент концентрации снаружи и внутри возбудимой клетки раздельно для ионов Na, К, Сl, и Са. Вопросы: Как изменится величина потенциала покоя и потенциала действия при увеличении градиента концентрации раздельно для ионов: 1) Na; 2) К; 3) Сl; 4) Сa?
1. При увеличении градиента концентрации Na величина потенциал покоя снизится, величина потенциала действия увеличится. 2. При увеличении градиента концентрации K величина потенциал покоя возрастет, величина потенциала действия не изменится. 3. При увеличении градиента концентрации Cl величина потенциал покоя возрастет, потенциала действия не изменится. 4. При увеличении градиента концентрации Са величина потенциал покоя снизится, величина потенциала действия не изменится. |
|
|
|
||
|
|
||
№ 10 Известно, что процесс возбуждения нервной и мышечной ткани характеризуется изменением ионной проницаемости клеточной мембраны. Экспериментально проводили избирательную блокаду мембранной проницаемости для ионов Na, К, Cl и Са в нервных волокнах и мышечных волокнах скелетной, гладкой и миокардиальной мышц. Вопрос: Как после этого изменится величина потенциалов покоя и действия в этих волокнах?
1. При блокаде мембранной проницаемости для Na величина потенциала покоя повысится, величина потенциала действия понизится. 2. При блокаде мембранной проницаемости для K величина потенциала покоя понизится, величина потенциала действия не изменится. 3. При блокаде мембранной проницаемости для K и для Cl величина потенциала покоя понизится, величина потенциала действия не изменится. 4. При блокаде мембранной проницаемости для K и для Ca величина потенциала покоя повысится, величина потенциала действия не изменится. В миокардиальных и гладкомышечных волокнах длительность потенциалов действия уменьшится в связи с ускорением реполяризации. |
№ 11 Известно, что суммация одиночных мышечных сокращений является одним из основных свойств мышечной ткани. В экспериментальных условиях изучали способность скелетной мышцы, отрезка кишки и сердца к суммации с помощью нанесения на эти органы двух последовательных раздражений. Вопросы: 1. Какие условия надо выполнить, чтобы добиться суммации одиночных сокращений? 2. Почему при суммации одиночных сокращений увеличивается амплитуда сокращения? 3. Какие виды мышц не способны к суммации одиночных сокращений, и почему это происходит? 4. При каких условиях повторяющиеся ритмические раздражения вызывают зубчатый тетанус, гладкий тетанус, оптимум и пессимум скелетной мышцы?
1. Для суммации одиночных сокращений необходимо соблюдение двух условий. Повторное раздражение должно наноситься во время первого одиночного сокращения и не должно попасть в период рефрактерности первого возбуждения. 2. Увеличение амплитуды сокращения при суммации двух одиночных сокращений связано с началом второго сокращения на фоне уже частично сократившейся мышцы. 3. Скелетная мышца способна к суммации одиночных сокращений. Длительность возбуждения и рефрактерного периода в скелетной мышце соответствует латентному периоду ее одиночного сокращения. Если повторное возбуждение попадает во время укорочения или расслабления первого одиночного сокращения, то второе сокращение суммируется с первым. В гладкой мышце длительность возбуждения и рефрактерный период соответствуют латентному периоду и фазе укорочения ее одиночного сокращения. Поэтому суммация двух сокращений может произойти в случае попадания второго раздражения в фазу расслабления гладкой мышцы. В сердце длительность возбуждения и рефрактерный период равны длительности одиночного сокращения. Поэтому суммация одиночных сокращений в миокарде не происходит. 4. Зубчатый тетанус возникает, когда каждое последующее раздражение попадает в фазу расслабления предыдущего одиночного сокращения. Гладкий тетанус возникает, когда каждое последующее раздражение попадает в фазу укорочения предыдущего одиночного сокращения. Оптимум возникает, когда каждое последующее раздражение попадает в фазу экзальтации предыдущего возбуждения. Пессимум возникает, когда каждое последующее раздражение попадает в абсолютный рефрактерный период предыдущего возбуждения. |
№ 12 Известно, что поддержание постоянства ионного состава в возбудимых тканях необходимо для их нормального функционирования. Отсутствие солей (и в частности — солей кальция) в питьевой воде у населения отдельного района России привело к нарушению функций скелетной мускулатуры. Обследование населения показало, что даже практически здоровые люди жаловались на повышенную мышечную утомляемость и недостаточную физическую силу. Вопросы: 1. Какова роль ионов кальция в механизме мышечного сокращения? 2. Почему недостаток кальция в организме сопровождается повышенной физической утомляемостью и недостаточной физической силой у людей? 3. Какие обследования целесообразности для изучения функций скелетной мускулатуры и миокарда?
1. В расслабленном мышечном волокне белок тропонин препятствует взаимодействию головок выростов миозина с актином. Мышечное сокращение начинается с выхода ионов кальция из Т-систем и эндоплазматического ретикулума. Ионы кальция соединяются с тропонином, и он сдвигается в сторону. Головки выростов миозина вступают в контакт с актином, обеспечивая скольжение нитей актина вдоль миозина. Длина саркомеров миофибрилл уменьшается. Длина всей мышцы уменьшается. 2. Количество связанных с тропонином ионов кальция определяет количество поперечных мостиков между нитями актина и миозина и, следовательно, силу и длительность сокращения. Поэтому недостаток в организме ионов кальция у людей приводит к уменьшению мышечной силы и повышению физической утомляемости. 3. Для изучения силы и выносливости скелетной мускулатуры необходимо исследовать динамометрию. Для изучения выносливости сердца необходимо провести исследование функций сердца с физическими нагрузками. |
|
|
|
||
|
|