Материал: Анализ соматических проявлений гибридного дисгенеза у Drosophila melanogaster (на модели признака cubitus interruptus Dominant)
Основными мерами защиты от поражения электрическим током являются:
обеспечение недоступности токопроводящих частей, которые находятся под напряжением, для случайного прикосновенья;
контроль и профилактика повреждений изоляции;
организация безопасной эксплуатации электроустановок.
Понятие пожарная безопасность означает состояние объекта, при котором исключается возможность пожара, а в случае ее возникновения предотвращается влияние на людей опасных факторов пожара и обеспечивается защита материальных ценностей.
В лаборатории используется электрооборудование, которое отвечает классу пожарной зоны помещения категории Д, степень огнестойкости III, степень защиты светильников не ниже ІР-2Х; категория защиты домов от молнии установлена ІІІ.
Согласно ГОСТ 12.1.004-91 [11] пожарная безопасность обеспечивается системой предотвращения пожара и пожарной защиты и проведением организационных мер. Помещение лаборатории оснащено углекислотным огнетушителем ВВК-1,4 и ведром с песком. Организационными мероприятиями по обеспечению пожарной безопасности является обучение работников кафедры правилам пожарной безопасности, разработка и реализация норм и правил пожарной безопасности, инструкций о порядке работы с пожароопасными материалами. Как профилактические меры по предупреждению причин пожаров используется постоянный контроль за состоянием электрической проводки и соединительных проводов. Обязательно наличие плана эвакуации. Ширина, высота и пропускная способность проходов и выходов отвечает противопожарным нормам.
Закон Украины об « Охране окружающей среды» был
принят 25 июня 1991 года (редакции Закона происходили в 1993 г., 1996 г.) [12].
Закон определяет правовые, экономические, социальные основы охраны окружающей
среды. Задача Закона состоит в регулировании отношений в области охраны
природы, использовании и воспроизведении природных ресурсов, обеспечении и
ликвидации последствий негативного влияния на окружающую среду хозяйственной и
другой деятельности человека, сохранение природных ресурсов, генетического
фонда наций, ландшафтов и других естественных объектов. Международные стандарты
включают требования сниженного энергопотребления и ограничения допустимых
уровней мощности, потребляемых в неактивном режиме. Необходимо выполнять
требования стандарта ISO - 14004 [16], что определяет требования к организации
производственного процесса с минимальным убытком, для окружающего природного
среды.
. РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЯ
. Анализ атрофии гонад
Анализ межполовых различий уровня атрофии гонад
в линиях и у гибридов показал следующее (Рис.4). Линия Пирятин характеризуется
высоким показателем АГ у самок, тогда как этот показатель у самцов данной линии
- в пределах нормы, что по данным литературы составляет около 5 % [27]. В
скрещивании ciDC-S x Пирятин уровень АГ у самцов не превышает допустимого
значения для данного вида, тогда как самки отличаются незначительным
увеличением этого показателя. Линия ciDC-S характеризуется достаточно высоким
показателем АГ, как у самцов, так и самок. У потомков от скрещивания ciDC-S x
W-10 выявлено, что уровень АГ самцов находится в пределах нормы, тогда как
самки характеризуются сравнительно высокими значениями данного показателя. В
линии W-10 более высокий уровень АГ установлен для самок и менее высокий - для
самцов.
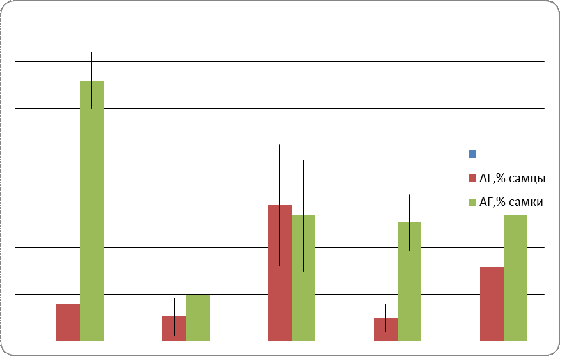
Рис. 4. Межполовые различия уровня атрофии гонад в линиях и у гибридов
При сравнении потомков от разных скрещиваний можно отметить, что самки в скрещивании ciDC-S x Пирятин характеризуются сниженным уровнем АГ по сравнению с контролем (линия Пирятин), то же самое наблюдается и с самцами, хотя в случае самцов снижение незначительно. Скрещивание ciDC-S x W-10 показало, что по сравнению с контролем (линия W-10) уровень АГ самцов снизился до уровня нормы. Этот показатель у самок тоже снизился, но незначительно и в целом остался на высоком уровне. При сравнении со вторым (линия ciDC-S) потомство от скрещивания ciD C-S x Пирятин характеризуется снижением уровня АГ как у самцов, так и у самок. В скрещивании ciDC-S x W-10 наблюдается незначительное уменьшение уровня АГ у самок, и резкое снижение (до уровня нормы) показателя у самцов.
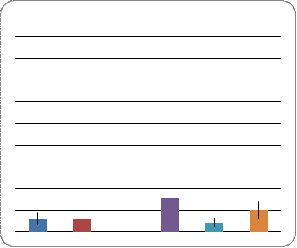
Экспрессивность признака ciD
Анализ экспрессивности признака ciD проводили у потомков двух гибридных скрещиваний (Рис. 5.), контролем для которых служила линия ciDC-S. Установлено, что в случае скрещивания ciDC-S x W-10 по сравнению с контролем у самок наблюдается незначительное уменьшение экспрессивности исследуемого признака, как на правом, так и на левом крыле. У самцов выявлены аналогичные изменения. При этом самцы - потомки скрещивания ciDC-S x W-10 характеризуются большей экспрессивностью признака ciD, нежели самки. Такой же результат был показан и для контроля линии ciDC-S, в которой большая экспрессивность ciD была характерна для самцов.
В скрещивании ciDC-S x Пирятин, по сравнению с контролем, экспрессивность ciD возросла у самцов и у самок на левом и правом крыле сравнительно одинаково. При этом у самок и самцов этот показатель находится примерно на одном уровне на обоих крыльях.
Во всех экспериментальных вариантах не выявлено
асимметрии в проявлении признака на левом и правом крыле у особей обоих полов.
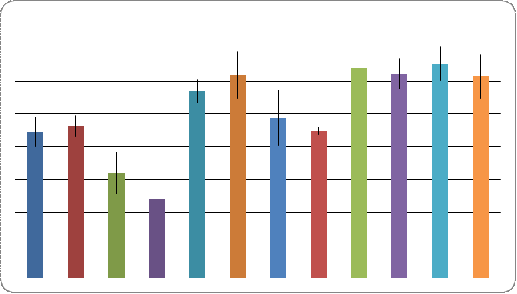
Рис. 5. Уровень экспрессивности признака ciD у
особей линии ciDC-S гибридов и у линии
Соотношение особей с атрофией гонад и наличием неполной 4-й жилки
Так как доминантный алелль ciD в гомозиготе летален, соответственно, выжившие особи с неполной 4-й жилкой являются гетерозиготами. Тогда в скрещиваниях гетерозигот и особей дикого типа ожидается два фенотипа потомков: в первом случае особи будут обладать выраженной экспрессивностью гена ciD и, соответственно, прерванной жилкой крыла; во втором - особи без мутантного фенотипа.
Все особи (потомки каждого скрещивания отдельно, самки и самцы отдельно) были разделены на 4 фенотипических класса (Рис. 6.):
Полная жилка, нормальные гонады
Полная жилка, атрофия гонад
Прерванная жилка, нормальные гонады
|
|
|
|
|
|
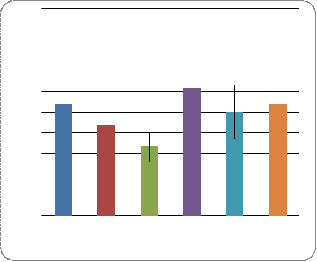 а) Полная
жилка, нормальные гонады
а) Полная
жилка, нормальные гонады
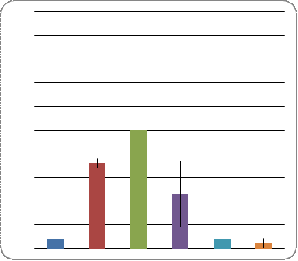 b) Полная
жилка. АГ
b) Полная
жилка. АГ
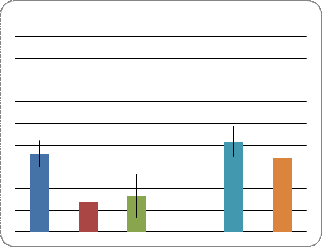 c)
Прерванная жилка, нормальные гонады
c)
Прерванная жилка, нормальные гонады
 d)
Прерванная жилка, АГ
d)
Прерванная жилка, АГ
Рис. 6. Соотношение особей с атрофией гонад и
наличием неполной 4-й жилки
Во всех вариантах скрещиваний 1-й класс численно преобладал, межполовых различий при этом не наблюдалось. Наименьшим количеством особей характеризовался 4-й класс. При этом в линии ciDC-S самцов с данным фенотипом не обнаружено. В двух других классах количество особей варьирует. Так, в скрещиваниях ciDC-S x W-10 и ciDC-S x Пирятин наибольшее количество особей мужского пола было с третьим фенотипом (прерванная жилка, нормальные гонады), тогда как со вторым фенотипическим классом (полная жилка, атрофия гонад) в данных скрещиваниях количество самцов приближалось к нулю. И наоборот, количество особей мужского пола во втором классе преобладало в линии ciDC-S. Самок этой линии во втором фенотипическом классе не обнаружено.
В скрещивании ciDC-S x Пирятин наименьшее количество особей соответствовало второму фенотипическому классу. Количество самок и самцов было минимально по сравнению с третьим классом, но приблизительно равным таковому в 4-м классе (прерванная жилка, атрофия гонад).
Полученные данные свидетельствуют о том, что экспрессивность признака ciD не связана напрямую с уровнем атрофии гонад. В формировании экспрессивности большое значение имеет генотип особи, линия.
Так, при уровне АГ до 10% у гибридов ciDC-S x Пирятин, экспрессивность ciD достигает высокого уровня и у самцов порядка 64%, и у самок до 65 %.
Линия ciDC-S характеризуется высокой экспрессивностью ciD (до 60 %). Гибриды от скрещивания её с линией W-10, содержащей полноразмерный Р-элемент, характеризуются снижением экспрессивности исследуемого признака, как у самцов (до 45 %), так и у самок (до 30 %). При этом уровень атрофии гонад у гибридных самцов также снижается, а у самок значимых изменений не выявлено.
Возможно, ожидаемого корреляционного эффекта не выявлено, так как оба показателя экспрессия ciD и уровень АГ противоположно зависят от температуры. Так, степень выраженности гонадной атрофии в отношении Р-М системы наиболее значительна при 29°С у самок и 27°С у самцов, а при 24° С и ниже практически отсутствует[30]. Для Н-Е системы характерна наиболее сильная атрофия при 25°С и наименьшая при 29° С[28]. При более низких температурах АГ наблюдается в меньшей степени. Экспериментальные скрещивания и линии содержались при 23°-24°С. Такая температура не случайна. Экспрессивность ciD при данной температуре достигает максимального значения, и с увеличением температуры до 25-26°С видимых проявлений мутации наблюдаться не будет.
Тем не менее, результаты скрещивания ciDC-S с линией содержащей полноразмерный hobo-элемент подтверждают, литературные данные, свидетельствующие о том, что индукция перемещений МГЭ может изменять проявление соматических мутаций[31], в данном случае - увеличивать экспрессивность признака ciD. Экспрессивность ciD у особей линии ciDC-S варьировала в пределах 58-61 % для самцов и до 50 % у самок, тогда как у гибридов ciDC-S x Пирятин экспрессивность достигла уровня 65% для самок и самцов.
Таким образом, несмотря на разнонаправленные
изменения экспрессивности признака ciD у гибридов разных дисгенных скрещиваний,
результаты проведенного исследования косвенно подтверждают литературные данные,
свидетельствующие о том, что индукция перемещения МГЭ в системах гибридного
дисгенеза может иметь соматические эффекты. Но для получения более точных
оценок необходимо провести несколько повторностей и проанализировать более
многочисленные выборки.
ВЫВОДЫ
Анализ литературных данных показал, что Р- и hobo -элементы вызывают не только синдром гибридного дисгенеза, но и существуют единичные данные, прямо или косвенно свидетельствующие о соматической активности этих элементов.
В ходе выполнения дипломной работы была освоена методика работы с D. melanogaster. Проанализированы линии и гибриды на предмет наличия синдрома гибридного дисгенеза, оценен уровень экспрессивности признака ciD.
Установлено, что у гибридов ciDC-S x Пирятин при
снижении уровня атрофии гонад наблюдается увеличение экспрессивности признака
ciD., а у гибридов ciDC-S x W-10 - при уменьшении уровня атрофии гонад -
снижение экспрессивности признака ciD.
СПИСОК ЛИТЕРАТУРЫ
Гинтер Е.К. Насекомые. В сб. «Методы биологии развития». // Ред. Детлаф Т.А. , «Наука»,- 1974, 272-281 с.
Голубовский М.Д. 1972. Некоторые общие характеристики организации генома и фенотипического проявления мутаций у дрозофилы. // Генетика - 1972.- Т. 8, № 5 - 143-157 с.
Антоненко О.В. Роль мобильных генетических элементов в отклике на селекцию по количественным признакам у Drosophila melanogaster. - Автореф. дис. канд. биол. наук. - Новосибирск ,- 2008. - 18 с.
Васильева Л.А. Изменение системы жилкования крыла Drosophila melanogaster под действием температурного шока и селекции // Журнал общей биологии. - 2005. - Т.66, №1. - С. 68-74.
ГОСТ 12.1.004-91 ССБТ. Пожарная безопасность. Общие требования. - Введ. 01.07.86; ГОСТ 12.1.004-91. ССБТ. Пожарная безопасность. Общие требования.- Чинний від 01.07.92.
ДНАОП - державний нормативний акт з охорони праці - 0.00 -4.12 - 99 «Типове положення про навчання з питань охорони праці»
ДСТ 12.0.003-74. ССБТ. Небезпечні і шкідливі виробничі фактории. Класифікація.
ДСТУ ISO 14004-97 Системи управління навколишнім середовищем. Загальні настанови щодо принципів управління, систем та засобів забезпечення. - Чинний від 18.08.97.
ЗАКОН УКРАЇНИ «Про державну систему біобезпеки при створенні, випробуванні, транспортуванні та використанні генетично модифікованих організмів». Документ 1103-16 <http://www.rada.gov.ua/cgi-bin/laws/main.cgi?nreg=1103-16>, остання редакцiя вiд 09.02.2010 <http://zakon1.rada.gov.ua/cgi-bin/laws/main.cgi?nreg=1103-16&c=1> на пiдставi 1804-17, чинний [http://zakon1.rada.gov.ua/cgi-bin/laws/main.cgi?nreg=1103-16].
Закон України «Про охорону навколишнього середовища» від 25.06.91.
Охорона праці (Законодавство. Організація роботи): Навч. посіб. / За заг. ред. к.т.н., доц. І. П. Пістуна. - Львів: “Тріада плюс”, 2010.- 648 с.
ПОСТАНОВА КМУ від 6 травня 2000 р. N 770 «Про затвердження переліку наркотичних засобів, психотропних речовин і прекурсорів» - Документ 770-2000-п <http://www.rada.gov.ua/cgi-bin/laws/main.cgi?nreg=770-2000-%EF>, остання редакцiя вiд 06.04.2011 <http://zakon1.rada.gov.ua/cgi-bin/laws/main.cgi?nreg=770-2000-%EF&c=1> на пiдставi 327-2011-п <javascript:OpenDoc('327-2011-п')>; <http://zakon1.rada.gov.ua/cgi-bin/laws/main.cgi?nreg=770-2000-%EF>
ПОСТАНОВА КМУвід 20 червня 1995 р. N 440 - «Про затвердження Порядку одержання дозволу на виробництво, зберігання, транспортування, використання, захоронення, знищення та утилізацію отруйних речовин, у тому числі продуктів біотехнології та інших біологічних агентів» - Документ 440-95-п <http://www.rada.gov.ua/cgi-bin/laws/main.cgi?nreg=440-95-%EF>, редакцiя вiд 06.01.2010 на пiдставi 1391-2009-п <javascript:OpenDoc('1391-2009-п')>; <http://zakon1.rada.gov.ua/cgi-bin/laws/main.cgi?nreg=440-95-%EF>
Правила устройства электроустановок. ПУЭ. Минэнерго СССР. - М.: Энергоатомиздат, 1987.
Про захист населення від інфекційних хвороб» - Документ 1645-14 <http://www.rada.gov.ua/cgi-bin/laws/main.cgi?nreg=1645-14>, остання редакцiя вiд 11.06.2009 <http://zakon1.rada.gov.ua/cgi-bin/laws/main.cgi?nreg=1645-14&c=1> на пiдставi 1254-17 <javascript:OpenDoc('1254-17')>, чинний; <http://zakon1.rada.gov.ua/cgi-bin/laws/main.cgi?nreg=1645-14>
«Про забезпечення санітарного та епідемічного благополуччя населення» - Документ 4004-12 <http://www.rada.gov.ua/cgi-bin/laws/main.cgi?nreg=4004-12>, поточна редакцiя вiд 12.03.2011 <http://zakon1.rada.gov.ua/cgi-bin/laws/main.cgi?nreg=4004-12&c=1> на пiдставi 3038-17 <javascript:OpenDoc('3038-17')>, чинний (Редакція вiдбудеться 12.06.2011 на пiдставi 3038-17 <javascript:OpenDoc('3038-17')>) <http://zakon1.rada.gov.ua/cgi-bin/laws/main.cgi?nreg=4004-12>://flybase.org/reports/FBgn0004859.html
Атраментова Л. О., Утєвськая О. М. Статистичні методи в біологіі: Підручник. - Х.: ХНУ ім. В. Н. Каразіна, 2007. - 288 с.
Жимулев И.Ф; под ред. Е.С. Беляева, А.П. Акифьева. Общая и молекулярная генетика - 4-е изд. - Новосибирск: Сиб. унив. изд-во, 2007. - 479 с.: ил.
Коваленко Л.В. hobo-элемент как фактор нестабильности генома Drosophila melanogaster в клетках генеративных и соматических тканей // Автореферат диссертации на соискание уч. степени к.б.н. -Российская академия наук. Сибирское отделение. Институт цитологии и генетики. - 2007. - 116 с.
Козерецкая И.А. Мобильные генетические элементы Drosophila melanogaster. Популяционно-генетический аспект.// Вісник Укр..тов-ва. генетиків і селекціонерів. - 2010.-Том 8, № 1.- 23 с.
Юшкова Е.А. Влияние хронического облучения в малых дозах на динамику изменчивости экспериментальных популяций Drosophila melanogaster, отличающихся по содержанию мобильных Р-элементов. // Автореферат диссертации на соискание уч.степени к.б.н. - Москва. - 2008. - 24 с.
Васильева Л.А., Захаров И.К. Селекция изменяет паттерн мобильных генетических элементов в геноме Drosophila melanogaster.// Вестник ВОГиС. - 2008. - Том 12, №3. - 412-424 с.
Вассерлауф И.Ф. Изменение организации хромосом в ядрах трофоцитов яичников Drosophila melanogaster при инбридинге и гибридном дисгенезе.// Вестник Томского государственного ун-та. - 2008. -. №316. - 178-186 с.
Голубовский М.Д. Нестабильность генов и мобильные элементы: к истории изучения и открытия.// Университет Калифорнии, Беркли, США; mdgolub@gmail.com. Историко-биологические исследования. - 2011. - Том 3, №4. - 60-76 с.
Буфф Е.М., Симонова О.Б. Участие мобильного элемента hobo в транспозицонных событиях в системе продлённой нестабильности у Drosophila melanogaster // Генетика. - 1992. - Т.28, №12. - 73-79 с.
Иващенко Н.И., Гришаева Т.М. Особенности индуцированного мутагенеза в системах гибридного дисгенеза у Drosophila melanogaster //Генетика.. - 2002. - Т. 38, №10. - 1351-1356 с.
Коваленко Л.В., Захаренко Л.П. Транспозиции hobo-элемента в соматических клетках Drosophila melanogaster // Генетика.. - 2006. - Т. 42., №2. - 177-184 с.
Юрченко Н.Н., Коваленко Л.В. Мобильные генетические элементы: нестабильность генов и геномов.// Вавиловский журнал генетики и селекции. - 2011.- Том 15, №2.- 261-266 с.
Кварец А.П., Мюссе Т.А. Динамика частоты гибридного дисгенеза у Drosophila melanogaster в контролируемых условиях хронического облученя.// Вестник Цитология и генетика.- 2010. - №3.- 21-26 с.