Материал: V-L-Bykov-Chastnaya_Gistologia_cheloveka-1
Колонка включает: 1) афферентные пути; 2) систему локальных связей и
3) эфферентные пути.
Афферентные пути. В центре колонки проходят примерно 100 возбуждающих кортико-кортикальных волокон - аксонов пирамидных клеток других колонок данного и противоположного полушарий. Они образуют окончания во всех слоях колонки (в том числе на клетках Мартинотти, шипиковых звездчатых клетках, латеральных дендритах пирамидных клеток) и проходят до I слоя, гае образуют ветви, уходящие за ее пределы.
Специфические афферентные импульсы по таламо-кортикальным волокнам поступают на тела и дендриты пирамидных клеток и на шипиковые звездчатые клетки IV слоя (последние по своим аксонам передают их на апикальные и базальные дендриты пирамидных клеток).
Система локальных связей формируется вставочными нейронами колонки,
которые включают более десятка типов клеток. Часть из них обладает тормозной функцией и регулирует преимущественно активность пирамидных клеток. Из тормозных нейронов колонки наибольшее значение имеют:
1)аксо-аксонные клетки, тела которых лежат во II-III слоях, а аксоны идут горизонтально, отдавая многочисленные терминальные веточки, которые образуют тормозные синапсы на начальных сегментах аксонов пирамидных клеток II и III слоев;
2)клетки-''канделябры" встречаются во всех внутренних слоях коры. Их аксонные коллатерали идут горизонтально и дают несколько восходящих и нисходящих веточек, которые образуют спиральные ветвления вокруг
апикальных дендритов пирамидных клеток. Описаны окончания аксонов клеток-«канделябров» на начальных сегментах аксонов пирамидных клеток (что сближает их с аксо-аксоннымн клетками);
3)корзинчатые клетки, которые располагаются во II слое, на границе III
иIV, а также IV и V слоев. Их аксоны проходят горизонтально на растояние до
2-3 мм и, оплетая тела крупных и средних пирамидных клеток, влияют на 20-30
соседних колонок. Колонковые корзинчатые клетки обеспечивают торможение пирамидных клеток по вертикали внутри данной колонки;
4)клетки с двойным букетам дендритов, отходящих вертикально от полюсов тела, расположенного II - III слоях. Их аксон дает коллатерали, образующие контакты с дендритами как пирамидных клеток, так и
непирамидных (в том числе тормозных) нейронов. Первый тип контактов опосредует угнетение пирамидных клеток, а второй - их активацию путем снятия торможения.
5)клетки с аксонным пучком (кисточкой) - звездчатые нейроны II слоя,
ахсоны которых ветвятся в I слое, образуя связи с дистальными сегментами апикальных дендритов пирамидных клеток и с горизонтальными ветвями
кортико-кортикальных волокон;
Эфферентные пути. Аксоны средних пирамидных клеток III слоя колонки устанавливают связи преимущественно с соседними колонками и колонками противоположного полушария, а аксоны крупных и гигантских пирамидных клеток V слоя, помимо этого, направляются в подкорковые центры, образуя вместе с аксонами веретеновидных клеток VI слоя систему эфферентных волокон коры.
Белое вещество головного мозга представлено пучками нервных волокон, которые поднимаются к серому веществу коры из ствола мозга и спускаются к стволу мозга от корковых центров серого вещества.
Глия головного мозга. Головной мозг содержит все виды макролии
(астроцитарную, эпендимную и олигодендроглию), а также микроглию.
Астроцитарная глия обеспечивает микроокружение нейронов, выполняет
опорную и трофическую функции в сером и белом веществе, участвует в метаболизме нейромедиаторов. Астроциты уплощенными пластинчатыми концевыми участками своих отростков образуют три вида пограничных глиальных мембран: периваскуллрные, поверхностную и субэпендимальную.
1) Пери васкулярные пограничные мембраны окружают капилляры головного мозга и входят в состав гемато-энцефалического барьера (ГЭБ),
отделяющего нейроны ЦНС от крови и тканей внутренней среды. ГЭБ препятствует проникновению в ЦНС переносимых кровью токсических веществ, нейромеднаторов. гормонов, антнбиоти-
ков (что затрудняет лечение инфекционных поражений мозга и его оболочек), поддерживает электролитный баланс мозга, обеспечивает избирательный транспорт ряда веществ (глюкозы, аминокислот) из крови в мозг.
Гамато-энцефалический барьер (ГЭБ) включает:
а) эндотелий кровеносных капилляров (с непрерывной выстилкой) -
главный компонент ГЭБ. Его клетки связаны мощными плотными соединениями, образование которых индуцируется контактом с астроцитами. Эндотелий препятствует переносу одних веществ, содержит специфические транспортные системы для других и метаболически изменяет третьи, превращая их в соединения, неспособные проникнуть в мозг (метаболический ГЭБ);
б) базальную мембрану капилляров, в) периеаскулярную пограничную глиальную мембрану из отростков
астроцитов.
2)Поверхностная пограничная глиальная мембрана (краевая глия) мозга,
расположена под мягкой мозговой оболочкой, образует наружную границу головного и спинного мозга, отдаляя ткани ЦНС от мозговых оболочек.
3)Субэпиндемальная (перивеитрнкулярная) пограничная глиальная мембрана располагается под слоем эпендимы и входит в состав нейроликворного барьера, который отделяет нейроны от спинномозговой жидкости (СМЖ), называемой также ликвором. Этот барьер представлен эпендимной глией, ее базальной мембраной (присутствует не везде) и отростками
астроципов.
Эпендимная глия образует выстилку желудочков головного мозга и входит в состав гемато-ликеорного барьера (между кровью и СМЖ).
Олигодендроглия встречается в сером и белом веществе; она обеспечивает
барьерную функцию, участвует в формировании миелиновых оболочек нервных волокон, регулирует метаболизм нейронов, захватывает нейромедиаторы.
Микроглия - специализированные макрофаги ЦНС, обладающие значительной подвижностью. Активируется при воспалительных и дегенеративных заболеваниях. Выполняют в ЦНС роль антиген-
представляющих дендритных клеток.
Желудочки головного мозга - система анастомозирующих полостей,
сообщающихся с центральным каналам спинного мозга и субарахноидальным пространством, содержащих СМЖ и выстланых однослойным пластом клеток эпендимной глии низкопризматической или кубической формы с микроворсинками и ресничками на апикальной поверхности. В отдельных участках эпендимоциты обладают специфическими структурнофункциональные особенностями и принимают участие в выработке СМЖ и химической сигнализации.
Сосудистые сплетения желудочков головного мозга - структуры в области крыши III и IV желудочков, а также части стенок боковых желудочков, которые обеспечивают выработку 70-90% СМЖ (10-30% вырабатываются тканями ЦНС и выделяются эпендимой вне области сосудистых сплетений). Они образованы ветвящимися выпячиваниями мягкой мозговой оболочки, которые вдаются в просвет желудочков и покрыты особыми кубическими хороидными эпендимоцитами (рис. 11-13).
Хороидные эпиндимоциты содержат большое количество митохондрий, умеренно развитый синтетический аппарат, многочисленные пузырьки и лизосомы. Их выпуклая апикальная поверхность покрыта многочисленными микроворсинками, латеральные формируют интердигитации и связаны комплексами соединений, а базальная образует переплетающиеся выросты (базальный лабиринт). По поверхности эпендимы сосудистых сплетений перемещаются уплощенные отростчатые клетки Кольмера с хорошо развитым лизосомальным аппаратом, которые, очевидно, являются макрофагами. Слой эпендимоцитов располагается на базальной мембране, отделяющей его от подлежащей рыхлой волокнистой соединительной ткани мягкой мозговой оболочки, в которой находятся многочисленные фенестрированные капилляры и встречаются слоистые обызвествленные тельца (конкрецин). Избирательная ультрафильтрация компонентов плазмы крови с образованием СМЖ происходит из капилляров в просвет желудочков через гемато-ликворный барьер. Установлено, что клетки эпендимы способны также секретировать некоторые белки в СМЖ и частично поглощать вещества из СМЖ (очищая ее от продуктов метаболизма мозга, лекарств, в частности, антибиотиков).
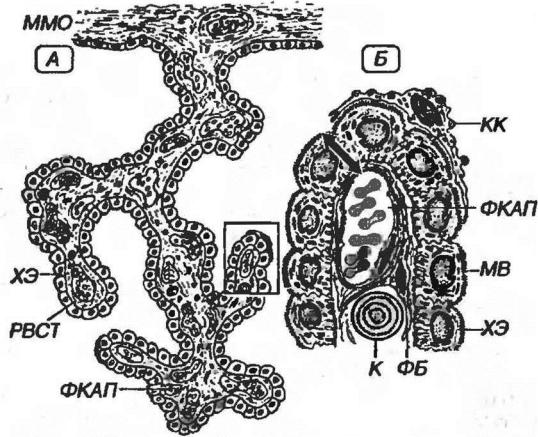
Гемато-ликворный барьер (см. рис. 11-13) включает:
1)цитоплазму фенестрированных эндотелиальных клеток капилляров,
2)базальную мембрану эндотелия капилляров,
3)перикапиллярное пространство - широкое, содержащее рыхлую волокнистую соединительную ткань мягкой мозговой оболочки с большим количестом макрофагов,
4)базальную мембрану эпендимы,
5)слой хороидных эпендимных клеток.
Рис. 11-13. Сосудистое сплетение желудочка головного мозга. А - общий вид участка сосудистого сплетения. ММО - мягкая мозговая оболочка, ХЭ - хориоидные эпендимоциты, РВСТ - рыхлая волокнистая соединительная ткань, ФКАП - фенестрированный капилляр. Е - фрагмент, выделенный рамкой на рисунка А: МВ - микро-ворсинки,ФБ - фибробласт, К - конкреция, КК - клетка Кольмера. Стрелкой показан гемато-ликворный барьер.
Спинномозговая жидкость циркулирует в субарахноидальном пространстве, желудочках головного мозга и центральной канале спинного мозга; ее общий объем у взрослого составляет 140-150 мл. Она вырабатывается в количестве 500 мл/сут, полностью обновляясь