Материал: V-L-Bykov-Chastnaya_Gistologia_cheloveka-1
Рис. 11-11. Цитоархитектоника (А и Б) и миелоархитектоника (В) коры полушарий большого мозга. Слои коры обозначены римскими цифрами. А - вид на препарате окрашенном гематоксилином и эозином; Б - вид не препарате, окрашенном методом серебрения по Гольджи; В - вид на препарате, окрашенном для избирательного выявления нервных волокон; ТВ - тангенциальные волокна, ПБ - полоски Бехтерева, НПБ - наружная полоска Байярже, ВПБ - внутренняя полоска Байярже, РЛ - радиальные лучи.
Непирамидные клетки располагаются практически во всех слоях коры, воспринимая поступающие афферентные сигналы, а их аксоны распространяются в пределах самой коры, передавая импульсы на пирамидные нейроны. Эти клетки весьма разнообразны и преимущественно являются разновидностями звездчатых клеток. Они включают шипиковые звездчатые, корзинчатые, аксо-аксонные клетки, клетки - "канделябры", клетки с двойным букетом дендритов, горизонтальные клетки Кахаля, клетки Мартинотти и
др. Основная функция не-пирамидных клеток - интеграция нейронных цепей внутри коры.
Цитоархитектоника коры полушарий большого мозга. Нейроны коры располагаются нерезко разграниченными слоями (пластинками), которые обозначаются римскими цифрами и нумеруются снаружи внутрь (см. рис. 11-11.
А, Б).
I - молекулярный слой располагается под мягкой мозговой оболочкой; содержит сравнительно небольшое число мелких нейронов - горизонтальных клеток Кахаля с длинными ветвящимися дендритами, отходящими в горизонтальной плоскости от веретеновидного тела. Их аксоны участвуют в образовании тангенциального сплетения волокон этого слоя. В молекулярном слое имеются многочисленные дендрит и аксоны клеток более глубоко расположенных слоев, образующих межнейронные связи.
II - наружный зернистый слой образован многочисленными мелкими
пирамидными и звездчатыми клетками, дендриты которых ветвится и поднимаются в молекулярный слой, в аксоны либо уходят в белое вещество, либо образуют дуги и также направляются в молекулярный слой.
III - пирамидный слой - значительно варьирует по ширине и максимально выражен в ассоциативных и сенсомоторных областях коры. В нем преобладают пирамидные клетки, размеры которых увеличиваются вглубь слоя от мелких до крупных. Апикальные дендрит пирамидных клеток направляются в молекулярный слой, а латеральные образуют синапсы с клетками данного слоя. Аксоны этих клеток оканчиваются в пределах серого вещества или направляются в белое. Помимо пирамидных клеток, слой содержит разнообразные непирамидные нейроны. Слой выполняет преимущественно ассоциативные функции, связывая клетки как в пределах данного полушария, так и с противоположным полушарием.
IV - внутренний зернистый слой - широкий в зрительной и слуховой областях коры, а в сенсомоторной области практически отсутствует. Он образован мелкими пирамидными и звездчатыми клетками. В этом слое заканчивается основная часть таламических афферентных волокон. Аксоны клеток этого слоя образуют связи с клетками выше- и нижележащих слоев коры.
V - ганглионарный слой образован крупными, а в области моторной коры (прецентральной извилины) - гигантскими пирамидными клетками (Беца). Апикальные дендриты пирамидных клеток достигают I слоя, образуя там верхушечные букеты, латеральные дендриты распространяются в пределах того же слоя. Аксоны гигантских и крупных пирамидных клеток проецируются на ядра головного и спинного мозга, наиболее длинные из них в составе пирамидных путей достигают каудальных сегментов спинного мозга. В V слое сосредоточено большинство корковых проекционных эфферентов.
VI - слой полиморфных клеток образован разнообразными по форме нейронами (веретеновидными, звездчатыми, клетками Мартинотти). Наружные участки слоя содержат более крупные клетки, внутренние - более мелкие и редко расположенные. Аксоны этих клеток уходят в белое вещество в составе эфферентных путей, а дендриты проникают до молекулярного слоя. Аксоны мелких клеток Мартинотти поднимаются к поверхности коры и ветвятся в молекулярном слое.
Миелоархитектоника коры полушарий большого мозга. Нервные волокна коры полушарий большого мозга включают три группы: 1) афферентные, 2)
ассоциативные и комиссуральные и 3) эфферентные волокна.
Афферентные волокна в виде пучков в составе радиальных лучей (см. рис. 11-11, В) приходят в кору от ниже расположенных отделов головного мозга, в частности, от зрительных бугров и коленчатых тел. Большая часть этих волокон заканчивается па уровне IV слоя.
Ассоциативные и комиссуральные волокна - внутрикорковые волокна,
которые соединяют между собой различные области коры в том же или в другом полушариях, соответственно. Эти волокна образуют пучки, которые проходят параллельно поверхности коры в I слое (тангенциальные волокна), во
II слое (полоска Бехтерева), в IV слое (наружная полоска Байярже) и в V слое
(внутренняя полоска Байярже). Последние две системы являются сплетениями, образованными конечными отделами афферентных волокон.
Эфферентные волокна связывают кору с подкорковыми образованиями. Эти волокна идут в нисходящем направлении в составе радиальных лучей (пример - пирамидные душ).
Типы строения коры. В отдельных участках коры, связанных с выполнением разных функций, преобладает развитие тех или иных ее слоев, на основании чего различают агранулярный и гранулярный типы коры.
Агранулярный тип коры характерен для ее моторных центров и отличается
наибольшим развитием III, V и VI слоев коры при слабом развитии II и IV
(зернистых) слоев. Такие участки коры служат источниками нисходящих проводящих путей ЦНС.
Гранулярный тип коры характерен для областей расположения чувствительных корковых центров. Он отличается слабым развитием слоев, содержащих пирамидные клетки, при значительной выраженности зернистых
(II и IV) слоев.
Нейромеккаторные системы коры. В коре выявлено более десятка различных
нейромедиаторов и рецепторов к ним, в том числе, норадреналин, серотонин, ацетилхолин, ВИП, соматостатин, холецистокинин, глутамат.
Модульный принцип организации коры полушарий большого мозга. В
коре полушарий большого мозга описаны повторяющиеся блоки (модули) нейронов, которые рассматривают как ее морфофункциональные единицы, способные к относительно автономной деятельности. Они имеют форму цилиндров, или колонок, диаметром 200-300 мкм (по некоторым данным, до 300 мкм и более), восходящих вертикально через всю толщу коры (рис. 11-12). В коре человека имеется около 2-3 млн. таких колонок, каждая содержит примерно 5000 нейронов. Внутри колонки выделяют также более мелкие миниколонки, включающие структуры, непосредственно окружающее апикальные дендрита пирамидных клеток.
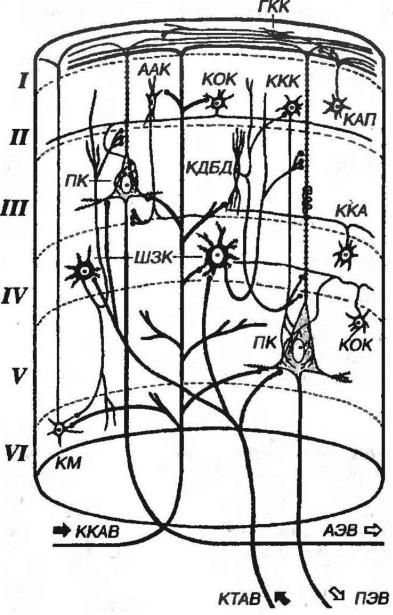
Рис. 11-12. Модуль (колонка) коры полушарий большого мозга. На схеме показаны главные ассоциативные связи клеток основных типов. Слои коры обозначены римскими цифрами. ККАВ - кортико-кортикальмое афферентное волокно, ТКАВ - таламо-кортикальное афферентное волокно, АЭВ - ассоциативное афферентное волокно, ПЭВ - проекционное афферентное волокно, ПК - пирамидное клетка, ШЗК - шипиковая звездчатая клетка. ГКК - горизонтальная клетка Кахаля. ААК – аксо-аксонная клетка, ККА - клетка- «канделябр», КОК - корзинчатая клетка, ККК - колонковая корзинчатая клетка, КДБД - клетка с двойным букетом дендритов, КАП - клетка с аксональным пучком, КМ - клетка Мартинотти. Пояснения в тексте.