Материал: V-L-Bykov-Chastnaya_Gistologia_cheloveka-1
нали преганглионарных волокон, стимуляйия которых приводит к усиленному выделению дофамина и других медиаторов в периваскулярные пространства и, возможно, в области синапсов на дендритах крупных клеток. МИФ-клетки обладают ингибирующим влиянием на активность эффекторных клеток.
Интрамуральные узлы и связанные с ними проводящие пути ввиду их высокой автономии, сложности оргаянизации и особенностей медиаторного обмена некоторыми авторами выделяются в самостоятельный
метасимпатичлский отдел вегетативной нервной системы. В частности,
общее число нейронов в интрамуральных узлах кишки выше, чем в спинном мозге, а по сложности их взаимодействия в регуляции перистальтики и секреции их сравнивают с миникомпьютером. Физиологически среди нейронов этих ганглиев выявлены клетки-водители ритма, которые обладают спонтанной активностью и посредством синаптической передачи воздействуют на "ведомые" нейроны, которые уже оказывают влияние на иннервируемые клетки.
Отсутствие части интрамуральных ганглиев толстой кишки вследствие дефекта их внутриутробного развития при врожденном заболевании (болезни Гиршпрунга) приводит к нарушению функции органа с резким расширением участка выше пораженного спазмированного сегмента.
В интрамуральных узлах описаны нейроны трех типов:
1)длинноаксонные эфферентные нейроны (клетки Догеля I типа)
численно преобладают. Это - крупные или средних размеров эфферентные нейроны с короткими дендритами и длинным аксоном, направляющимся за пределы узла к рабочему органу, на клетках которого он образует двигательные или секреторные окончания (см. рис. 11-4. А);
2)равноотростчатые афферентные нейроны (клетки Догеля II типа)
содержат длинные дендриты и аксон, уходящий за пределы данного ганглия в соседние и образующий синапсы на клетках I и III типов. Эти клетки, повидимому, входят в качестве рецепторного эвена в состав местных рефлекторных дуг, которые замыкаются без захода нервного импульса в ЦНС. Наличие таких дуг подтверждается сохранением функционально активных афферентных, ассоциативных и эфферентных нейронов в трансплантированных органах (например, сердце);
3) ассоциативные клетки (клетки Догеля III типа) - местные
вставочные нейроны, соединяющие своими отростками несколько клеток I и II типов, морфологически сходные с клетками Догеля II типа. Дендрита этих клеток не выходят за пределы узла, а аксоны направляются в другие узлы, образуя синапсы на клетках I типа.
СПИННОЙ МОЗГ
Спинной мозг располагается в позвоночном канале и имеет вид округлого тяжа, расширенного в шейном и поясничном отделах и пронизанного центральным каналом. Он состоит из двух симметричных половин,
разделенных спереди срединной щелью, сзади - срединной бороздой, и
характеризуется сегментарным строением; с каждым сегментом связана пара передних (вентральных) и пара задних (дорсальных) корешков. В спинном мозге различают серое вещество, расположенное в его центральной части, и белое вещество, лежащее по периферии (рис. 11-5).
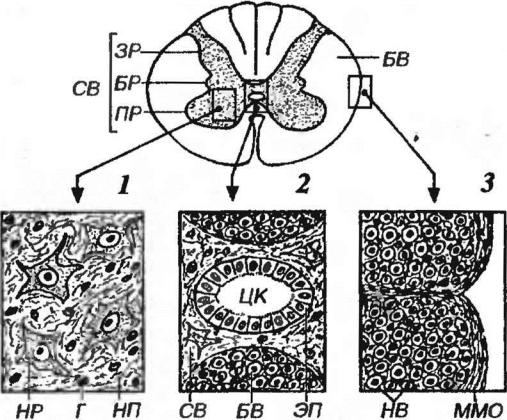
Рис. 11-5. Строение различных участков спинного мозга. СВ - серое вещество, ЗР - задние рога, БР - боковые рога, ПР - передние рога, БВ - белое вещество. 1 -серое вещество: НР - нейрон (тело), Г - глиоцит (ядро), НП – нейропиль. 2 - область центральной серой комиссуры с центральным каналом (ЦК), ЭП - эпендима. 3 - белое вещество: НВ - нервные волокна, ММО - мягкая мозговая оболочка.
Серое вещество на поперечной разрезе имеет вид бабочки (см. рис. 11-5, 1) и
включает парные передние (вентральные), задние (дорсальные) и боковые
(латеральные) рога (в действительности представляют собой непрерывные столбы, идущие вдоль спинного мозга). Рога серого вещества обеих симметричных частей спинного мозга связаны друг с другом в области
центральной серой комиссуры (спайки). В сером веществе находятся тела,
дендриты и (частично) аксоны нейронов, а также глиальные клетки. Между телами нейронов находится нейропиль - сеть, образованная нервными волокнами н отростками глиальных клеток.
Цнтоархитектоника спинного мозга. Нейроны располагаются в сером веществе в виде не всегда резко разграниченных скоплений (ядер), в которых происходит переключение нервных импульсов с клетки на клетку (отчего их относит к нервным центрам ядерного типа). На основании расположения нейронов, их цитологических особенностей, характера связей и функции. Б.Рекседом в сером веществе спинного мозга выделены десять пластин, идущих в ростро-каудальном направлении. В зависимости от топографии аксонов нейроны спинного мозга подразделяют на: 1) корешковые нейроны, аксоны которых образуют передние корешки; 2) внутренние нейроны, отростки которых заканчиваются в пределах серого вещества спинного мозга; 3) пучковые нейроны, отростки которых образуют пучки волокон в белом веществе спинного мозга в составе проводящих путей.
Задние рога содержат несколько ядер, образованных мультиполярными вставочными нейронами мелких в средних размеров, на которых оканчиваются аксоны псевдоунмиполярных клеток спинальных гангиев, несущие разнообразную информацию от рецепторов, а также волокна нисходящих путей и лежащих выше (супраспинальных) центров. В задних рогах выявляются высокие концентрации таких нейромедиаторов, как серотонин, энкефалин,
вещество Р.
Аксоны вставочных нейронов а) оканчиваются в сером веществе спинного мозга на мотонейронах, лежащих в передних рогах; б) образуют межсегментарные связи в пределах серого вещества сшитого мозга; в) выходят в белое вещество спинного мозга, где образуют восходящие и нисходящие проводящие пути (тракты). Часть аксонов при этом переходит на противоположную сторону спинного мозга.
Боковые рога, хорошо выраженные на уровне грудных и крестцовых сегментов спинного мозга, содержат ядра, образованные телами вставочных нейронов, которые относятся к симпатическому и парасимпатическому отделам вегетативной нервной системы (см. рис. 11-1, А). На дендритах в телах этих клеток оканчиваются аксоны: а) псевдоуниполярных нейронов, несущих импульсы от рецепторов, расположенных во внутренних органах, б) нейронов центров регуляция вегетативных функций, тела которых находятся в продолговатом мозге. Аксоны вегетативных нейронов, выходя из спинного мозга в составе передних корешков, образуют преганглионарные волокна,
направляющиеся к симпатическим и парасимпатическим узлам. В нейронах боковых рогов основным медиатором является ацетилхолин, выявляется также ряд нейропептидов - энкефолин, нейротензин, ВИП, вещество Р, соматостатин, пептид, связанный с кальцитониновым геном (ПСКГ).
Передние рога содержат мультиполярные двигательные клетки
(мотонейроны) общим числом около 2-3 млн. Мотонейроны объединены в ядра, каждое из которых обычно тянется на несколько сегментов. Различают крупные (диаметр тела 35-70 мкм) альфа-мотонейроны и рассеянные среди них более мелкие (15-35 мкм) гамма-мотонейроны.
На отростках и телах мотонейронов имеются многочисленные синапсы (до нескольких десятков тысяч на каждом), оказывающие на них возбуждающие и тормозные воздействия. На мотонейронах оканчиваются:
а) коллатерали аксонов псевдоуниполярных клеток спинальных узлов, образующие с ними двухнейронные (моносинаптические) рефлекторные дуги (см. рни. 11-1 и рис. 11-6);
б) аксоны вставочных нейронов, тела которых лежат в задних рогах спинного мозга;
в) аксоны клеток Реншоу, образующие тормозные аксо-соматнческие синапсы (см. рис. 11-6). Тела этих мелких вставочных ГАМК-эргических нейронов располагаются в середине переднего рога и иннервированы коллатералями аксонов мотонейронов,
г) волокна нисходящих путей пирамидной и экстрапирамндной систем, несущие импульсы из коры большого мозга и ядер ствола мозга.
Гамма-мотонейроны, в отличие от альфа-мотонейронов, не имеют непосредственной связи с чувствительными нейронами спинальных узлов.
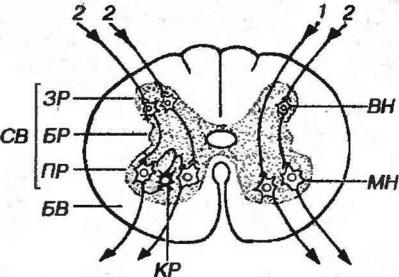
Рис. 11-6. Основные ассоциативные связи нейронов в сегменте спинного мозга. 1 - моносимаптческая (двухнейронная) рефлекторная дуга, 2 - дисинаптическая (трехнейронная) рефлекторная дуга. СВ - серое вещество, ЗР - задние рога, БР - боковые рога, ПР - передние рога, БВ - белое вещество, ВН - вставочный нейрон, МН - мотонейрон КР - клетка Реншоу.
Аксоны альфа-мотонейронов отдают коллатерали, оканчивающиеся на телах вставочных клеток Реншоу (см. выше), и покидают спинной мозг в составе
передних корешков, направляясь в смешанных нервах к соматическим мышцам, на которых они заканчиваются нервно-мышечными синапсами (моторными бляшками). Более тонкие аксоны гамма-мотонейгронов имеют такой же ход и образуют окончания на интрафузальных волокнах нервно мышечных веретен. Нейромедиатором клеток передних рогов является ацетилхолин.
Центральный (спинномозговой) канал (см. рис. 11-5, 2) проходит в центре серого вещества в центральной серой комиссуре (спайке). Он заполнен
спинномозговой жидкостью (СМЖ) и выстлан одним слоем кубических или призматических клеток эпендимы, апикальная поверхность которых покрыта микроворсинками и (частично) ресничками, а латеральные связаны комплексами межклеточных соединений.
Белое вещество спинного мозга окружает серое (см. рис. 11-5, 3) и разделяется передними и задними корешками на симметричные дорсальные,
латеральные и вентральные канатики. Оно состоит из продольно идущих нервных волокон (преимущественно миелиновых), образующих нисходящие и восходящие проводящие пути (тракты). Последние отделены друг от друга тонкими прослойками соединительной