Материал: Obmen_i_funktsii_aminokislot_Osnovy_molekulyarnoi_774_genetiki
• РНК-полимераза I, синтезирующая пре-рРНК;
|
• РНК-полимераза II, ответственная за синтез пре-мРНК; |
• РНК-полимераза III, синтезирующая пре-тРНК.
Синтез начинается с того момента, когда РНК-полимераза II присоединяется к матрице в специальном участке - протомотре. Промотор содержит ТАТА-последоательность, которую узнают белком ТАТА-фактором. Матрицей для синтеза служит одна из цепей ДНК.
Элонгация - наращивание молекулы РНК происходит путем присоединения очередного рибонуклеотида, комплементарного тому дезоксирибонуклеотиду ДНК, который находится в активном центре РНК-полимеразы
Терминация. участок ДНК содержит определенный участок - сайт терминации, где заканчивается ген, представляющий из себя последовательность нуклеотидов.И фермент отделяется ДНК-цепь.
Модификации пре-мРНК начинаются на стадии элонгации. Когда длина первичного транскрипта достигает примерно 30 нуклеотидов, происходит «кэпирование» 5'-конца.
Первичный транскрипт или пре-мРНК комплементарен гену, содержит как экзоны - последовательности, кодирующие определенные участки молекулы белка, так и интроны - некодирующие последовательности. В процессе образования молекул «зрелой» мРНК интроны вырезаются из первичного транскрипта, концы экзонов соединяются друг с другом - эту реакцию называют сплайсингом РНК
Процесс вырезания интронов протекает при участии малых ядерных рибонуклеопротеинов (мяРНП), которые образуют комплексы - сплайсосомы. После завершения сплайсинга «зрелая»
мРНК становится примерно в четыре раза короче первичного транскрипта. Сплайсинг происходит в ядре, в цитоплазму переносится уже «зрелая» мРНК
66. Понятие о биологическом коде, свойства биологического кода.
Универсальность биологического кода и процессов биосинтеза белка.
Биологический код - это способ записи информации об аминокислотной последовательности белков с помощью последовательности нуклеотидов в ДНК или РНК. Его характеризуют следующие свойства: триплетность и наличие терминирующих кодонов, специфичность, вырожденность, универсальность, однонаправленность, колинеарность
|
Триплетность и наличие терминирующих кодонов |
Кодовое число равно 3. Три нуклеотидных остатка (триплет) кодируют одну аминокислоту. Терминирующие триплеты - UАА, UАG, UGА не кодируют аминокислот, а являются сигналами к прекращению синтеза белка |
|
Специфичность |
Каждый триплет кодирует только одну аминокислоту |
|
Вырожденность |
Одну аминокислоту могут кодировать несколько триплетов (от 2 до 6) |
|
Универсальность |
Почти у всех видов организмов биологический код одинаков |
|
Однонаправленность |
Информация, записанная в зрелой мРНК в виде линейной последовательности кодонов (триплетов), считывается в направлении от 5'- к 3'-концу |
|
Колинеарность |
Последовательность кодонов в зрелой мРНК соответствует последовательности аминокислот в синтезированном белке |
67. Транспортная рнк как адаптор аминокислот. Биосинтез аминоацил-т-рнк.
Второй тип РНК называют транспортной (адапторной сокращенно— т-РНК). Она выполняет функцию переносчика активированных аминокислотк месту синтеза белка. Каждая молекула такой РНК является адаптером только какой-нибудь одной аминокислоты, т. е. находит место аминокислоты на и-РНК- Поэтому в клетке имеется не менее 20 разновидностей РНК, специализированных для 20 аминокислот.
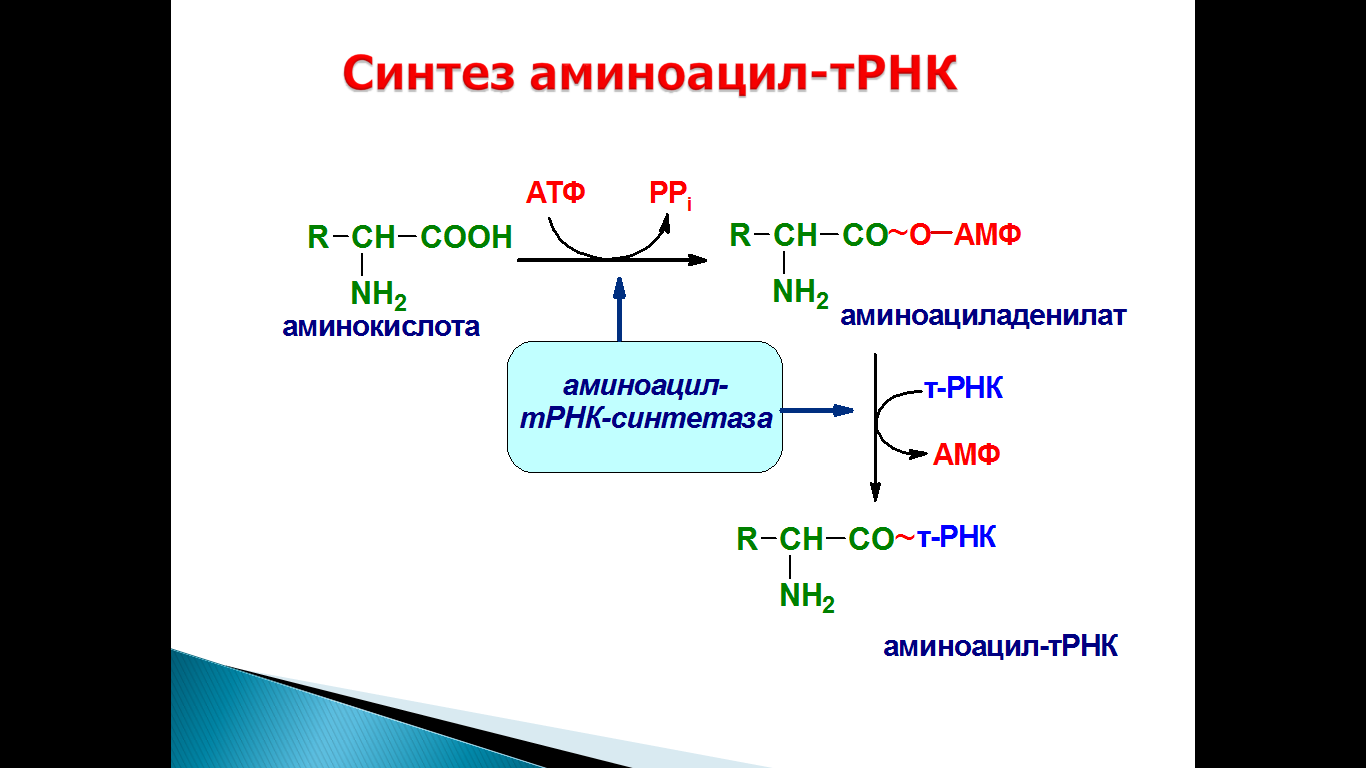
68. Субстратная специфичность арс-аз, их роль. Изоакцепторные т-рнк.
Реакция аминоацилирования осуществляется ферментами аминоацил-тРНК-синтетазами, способными узнавать три различных субстрата: АТФ, аминокислоту и тРНК. В активном центре молекулы фермента осуществляются активация аминокислоты и присоединение ее к концевому остатку рибозы тРНК. Для каждой аминокислоты в клетке имеется аминоацил-тРНК-синтетаза (АРСаза), способная отличать свою аминокислоту от остальных, часто очень похожих по структуре/
Аминоацил-тРНК-синтетазы осуществляют присоединение аминокислоты к молекуле транспортной РНК, что является ключевым моментом в реализации генетической информации.
Изоакцепторные тРНК. - Группа тРНК, связывающих одну и ту же аминокислоту, но имеющих разные антикодоны; разные Изоакцепторные тРНК узнаются одной и той же аминоацил-тРНК-синтетазой; Изоакцепторные тРНК отсутствуют у метионина и триптофана, а наибольшее их число (по 6) распознают кодоны аденина, лейцина и серина;Изоакцепторные тРНК могут иметь одинаковые антикодоны, но различную первичную структуру.
69. Строение рибосом. Последовательность событий на рибосоме при сборке полипептидной цепи, функционирование полирибосом.
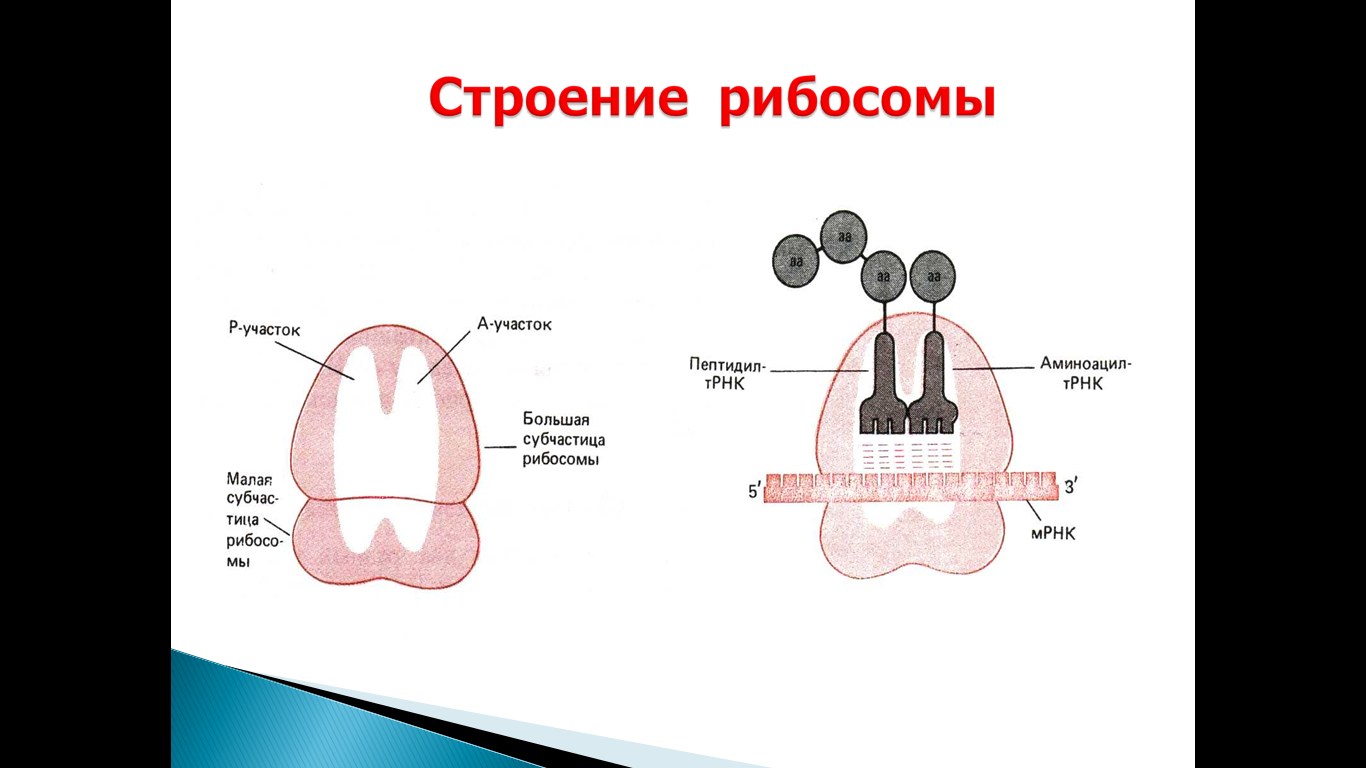
События на рибосоме включают этапы: инициации, элонгации и терминации.
Инициация начинается с присоединения к мРНК в области «кэпа» малой субъединицы рибосомы 40S, факторов инициации (IF), инициирующей Мет-тРНКМет и ГТФ. Когда в результате движения этого комплекса по мРНК антикодон Мет-тРНКМет свяжется с инициирующим кодоном АUG, комплекс останавливается. Происходит присоединение 60S-субъединицы рибосомы, сопровождающееся гидролизом ГТФ и отделением факторов инициации. Формируется 80S-рибосома с двумя активными центрами: Р (пептидильным) центром, в котором находится Мет-тРНКМет, и А(аминоацильным) центром, в область которого поступает первый смысловой кодон
|
|
мРНК.
Этап элонгации включает три последовательные стадии:
- Связывание аа-тРНКаа в А-центре.
- Образование пептидной Пептидил-тРНК связи.
- Транслокация - перемещение рибосомы по мРНК.
Терминация трансляции происходит после включения в А-центр одного из стоп кодонов: UAG, UGA, UAA
Одновременно несколько рибосом могут участвовать в трансляции одной мРНК. Каждая рибосома занимает участок, равный примерно 80 нуклеотидам мРНК. Таким образом, рибосомы располагаются на мРНК с интервалами около 100 нуклеотидов, образуя комплекс, называемый полисомой.
70. Посттрансляционный процессинг белков.
Функционально активные белки образуются в результате посттрансляционных модификаций полипептидных цепей, синтезированных на рибосомах. Они включают:
• частичный протеолиз;
• фолдинг, или формирование пространственной структуры, в котором принимают участие белки-шапероны, обеспечивающие образование функционально активной конформации полипептидной цепи;
• модификации аминокислот: карбоксилирование, фосфорилирование, йодирование, гидроксилирование, ацилирование и гликозилирование;
• образование дисульфидных связей между остатками цистеина, участвующими в формировании трехмерной структуры белка;
• присоединение простетических групп;
• образование олигомерных структур, которое также осуществляется при участии шаперонов.
71. Адаптивная регуляция экспрессии генов у прокариотов и эукариотов.
Адаптивная регуляция активности генов у прокариотов получила объяснение в теории оперона. Согласно этой теории на молекуле ДНК прокариотов присутствуют определенные участки - опероны. В состав этих участков ДНК входятструктурные гены, содержащие информацию о группе функционально взаимосвязанных белков, которые участвуют в одном и том же метаболическом пути, промотор и оператор. Участки промотора и оператора частично перекрываются. Транскрипцию структурных генов контролирует оператор, присоединение к которому белка-репрессора не позволяет РНК-полимеразе связаться с промотором и начать транскрипцию. Белок-репрессор синтезируется в клетке с постоянной скоростью, его строение кодирует мРНК, транскрибируемая с гена-регулятора, расположенного на некотором расстоянии от оперона, работу которого контролирует его белковый продукт.
Адаптивная регуляция активности генов у эукариотов обеспечивает изменения скорости транскрипции отдельных генов в ответ на меняющиеся условия внутренней и внешней среды. В клетках многоклеточных организмов часть генов кодирует белки «домашнего хозяйства», которые синтезируются с постоянной скоростью и обеспечивают жизнеспособность клеток. Это - гены ферментов, участвующие в биологическом окислении, синтезе АТФ, образовании компонентов мембран и т.д.
|
|
Регуляция у высших организмов отличается от регуляции транскрипции у прокариотов многообразием сигналов, которые контролируют не только начало процесса на молекуле ДНК, но и частоту, с которой он происходит.