Материал: 59-64
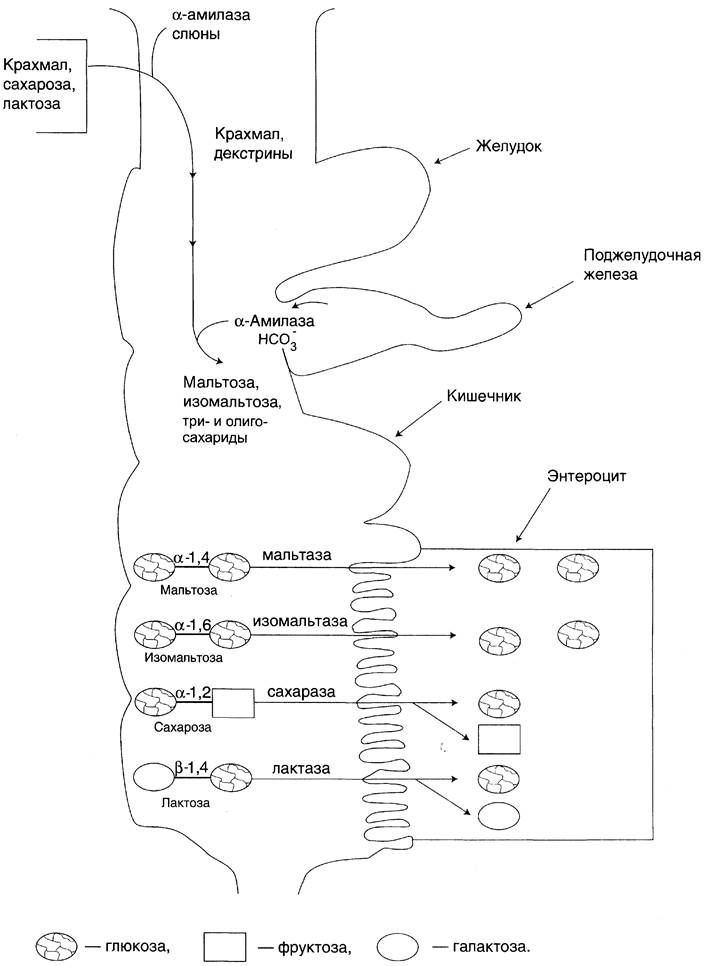
2. Путем облегченной диффузии – когда концентрация глюкозы в просвете
кишечника велика
После моносахариды покидают клетки слизистой оболочки кишечника с помощью
облегчённой диффузии. Часть глюкозы (более 50%) через капилляры кишечных ворсинок попадает в кровеносную систему и по воротной вене доставляется в печень. Остальное количество глюкозы поступает в клетки других тканей.
Г. Транспорт глюкозы из крови в клетки: потребление глюкозы клетками из кровотока происходит путём облегчённой диффузии – получается, что скорость трансмембранного потока глюкозы зависит только от градиента её концентрации.
Исключение: клетки мышц и жировой ткани, где облегчённая диффузия
регулируется инсулином (гормон поджелудочной железы). В отсутствие инсулина плазматическая мембрана этих клеток непроницаема для глюкозы, так как она не содержит белки-переносчики (транспортёры) глюкозы.
Способ облегчённой диффузии по сравнению с активным транспортом предотвращает транспорт ионов вместе с глюкозой, если она транспортируется по градиенту концентрации.
Глюкозные транспортёры (ГЛЮТ) обнаружены во всех тканях, существует
несколько их разновидностей. Структура белков семейства ГЛЮТ отличается от белков, транспортирующих глюкозу через мембрану в кишечнике и почках против градиента концентрации.

Все типы ГЛЮТ могут находиться как в плазматической мембране, так и в
цитозольных везикулах. ГЛЮТ-4 (и в меньшей мере ГЛЮТ-1) почти полностью находятся в цитоплазме клеток.
Влияние инсулина на такие клетки: приводит к перемещению везикул, содержащих
ГЛЮТ, к плазматической мембране, слиянию с ней и встраиванию транспортёров в мембрану = облегчённый транспорт глюкозы в эти клетки. После снижения конц. инсулина в крови ГЛЮТ снова перемещаются в цитоплазму, и поступление глюкозы в клетку прекращается.
Перемещение глюкозы из первичной мочи в клетки почечных канальцев
происходит вторично-активным транспортом (как и при всасывании глюкозы из просвета кишечника в энтероциты). Благодаря этому глюкоза может поступать в клетки даже в том случае, если её концентрация в первичной моче меньше, чем в клетках. При этом глюкоза реабсорбируется из первичной мочи почти полностью (99%).
Вопрос 62
Глюкоза как важный метаболит углеводного обмена: общая схема источников и путей расходования глюкозы в организме. Поддерживание постоянного уровня глюкозы крови, количественное определение глюкозы крови.
В глюкозу превращаются углеводы в печени и из глюкозы могут образовываться
все остальные углеводы. Т. о., она играет роль связующего между энергетическими и пластическими функциями углеводов.
Основные источники глюкозы:
Пища
Распад резервного полисахарида гликогена
Синтез глюкозы из неуглеводных предшественников
(главным образом из гликогенных аминокислот)
Глюконеогенез
Основные пути расходования глюкозы:
Образование энергии при аэробном и анаэробном окислении глюкозы
Превращение в другие моносахариды
Превращение в гликоген и гетерополисахариды
Превращение в жир, некоторые аминокислоты и др.
Уровень глюкозы в организме постоянный. Если глюкоза не поступает с пищей, то у здорового человека в крови поддерживается постоянная концентрация глюкозы от 70 до 90 мг/100 мл. После приема содержащей углеводы пищи концентрация глюкозы на короткое время возрастает примерно до 150 мг/ 100 мл, но через 2 ч она обычно возвращается к норме.
После всасывания в кишечнике моносахариды поступают в воротную вену и далее
преимущественно в печень. Поскольку в составе основных углеводов пищи преобладает глюкоза = она осн. продукт переваривания углеводов. Другие моносахариды, поступающие из кишечника в процессе метаболизма, могут превращаться в глюкозу или продукты её метаболизма.
Часть глюкозы в печени депонируется в виде гликогена, а другая часть через
общий кровоток доставляется и используется разными тканями и органами.
Фосфорилирование: глюкоза, поступающая в клетки органов и тканей, сразу же
подвергается фосфорилированию с использованием АТФ.
Эту реакцию во многих тканях катализирует гексокиназа,
а в печени и поджелудочной железе – фермент глюкокиназа.
Образование глюкозо-6-фосфата в клетке – своеобразная «ловушка» для глюкозы,
так как мембрана клетки непроницаема для фосфорилированной глюкозы (нет соответств. транспортных белков). Кроме того, фосфорилирование уменьшает конц. свободной глюкозы в цитоплазме. В результате создаются благоприятные условия для облегчённой диффузии глюкозы в клетки из крови.
Глюкокиназа. В период пищеварения конц. глюкозы в воротной венебольше, чем в
других отделах кровяного русла = активность глюкокиназы в гепатоцитах повышается.
Активность глюкокиназы, в отличие от гексокиназы, не ингибируется продуктом
(глюкозо-6-фосфатом), что обеспечивает повышение концентрации глюкозы в клетке в фосфорилированной форме, соответственно её уровню в крови.
Глюкоза проникает в гепатоциты путём облегчённой диффузии при участии
транспортёра ГЛЮТ-2 (независимого от инсулина).
Хотя инсулин и не влияет на транспорт глюкозы, он усиливает приток
глюкозы в гепатоциты в период пищеварения косвенным путём, индуцируя синтез глюкокиназы и ускоряя тем самым фосфорилирование глюкозы.
Преимущественное потребление глюкозы гепатоцитами, обусловленное
свойствами глюкокиназы, предотвращает чрезмерное повышение её концентрации в крови в абсорбтивном периоде – это снижает последствия протекания нежелательных реакций с участием глюкозы, например, гликозилирования белков.
Гексокиназа отличается от глюкокиназы высоким сродством к глюкозе –
он, в отличие от глюкокиназы, активен при низкой концентрации глюкозы в крови, что характерно для постабсорбтивного состояния.
Печень в этот период поглощает гораздо меньше глюкозы, так как скорость её
внутриклеточного фосфорилирования глюкокиназой резко снижается. Тогда как потребление глюкозы мозгом, эритроцитами и другими тканями обеспечивается активной в этих условиях гексокиназой.
Активность гексокиназы изменяется в зависимости от потребностей клетки в
энергии. В качестве регуляторов выступают соотношение АТФ/АДФ и внутриклеточный уровень глюкозо-6-фосфата. При снижении расхода энергии в клетке повышается уровень АТФ (относительно АДФ) и глюкозо-6-фосфата = активность гексокиназы снижается и уменьшается скорость поступления глюкозы в клетку.
Дефосфорилирование глюкозо-6-фосфата: превращение глюкозо-6-фосфата в
глюкозу возможно в печени, почках и клетках эпителия кишечника.
В них есть глюкозо-6-фосфатаза, катализирующая отщепление фосфатной группы гидролитическим путём:
Глюкозо-6-фосфат +Н2O → Глюкоза + Н3РO4
Образовавшаяся свободная глюкоза способна диффундировать из этих органов в кровь.
В других органах и тканях глюкозо-6-фосфатазы нет, и поэтому
дефосфорилирование глюкозо-6-фосфата невозможно (например, в мышцах)
Метаболизм глюкозо-6-фосфата: он может использоваться в клетке в различных
превращениях, основными из которых являются: синтез гликогена, катаболизм с образованием СО2 и Н2О или лактата, синтез пентоз
Поддержание уровня глюкозы в крови: распад гликогена печени служит в
основном для поддержания уровня глюкозы в крови в постабсорбтивном периоде.
Гликоген депонируется в печени и скелетных мышцах.
Содержание гликогена в печени изменяется в зависимости от ритма питания.
При длительном голодании оно снижается почти до нуля.
Гликоген мышц служит резервом глюкозы – источника энергии при мышечном
сокращении. Мышечный гликоген не используется для поддержания уровня глюкозы в крови (в клетках мышц нет фермента глюкозо-6-фосфатазы, и образование свободной глюкозы невозможен), так что его расход зависит в основном от физической нагрузки
Определение глюкозы в крови – важный этап диагностики сахарного диабета.
Для этого необходимо определить содержание глюкозы в крови. В норме ее конц. лежит в пределах 3,3-5,5 ммоль/л.
Методики определения: редуктометрические, колориметрические, ферментативные
Ферментативные методы определения конц. глюкозы в крови наиболее
распространены. Выделяют две основные разновидности этих методов: глюкозооксидазный и гексокиназный. На данный момент наиболее распространенными являются глюкозооксидазные: этот фермент реагирует с глюкозой, в результате чего образуется перекись водорода. Количество образовавшейся перекиси водорода равно количеству глюкозы в исходной пробе.
Вопрос 63 Аэробный гликолиз. Последовательность реакций до образования пирувата (аэробный гликолиз). Физиологическое значение аэробного гликолиза. Использование глюкозы для синтеза жиров.
Аэробный гликолиз – процесс окисления глюкозы до пировиноградной
Кислоты (ПВК), протекающий в присутствии кислорода. Все ферменты, катализирующие реакции этого процесса, локализованы в цитозоле клетки.
Этапы аэробного гликолиза: можно выделить 2 этапа
Подготовительный этап, в ходе которого глюкоза фосфорилируется и
расщепляется на две молекулы фосфотриоз. Эта серия реакций протекает с использованием 2 молекул АТФ.
Этап, сопряжённый с синтезом АТФ. В результате этой серии реакций
фосфотриозы превращаются в пируват. Энергия, высвобождающаяся на этом этапе, используется для синтеза 10 моль АТФ.
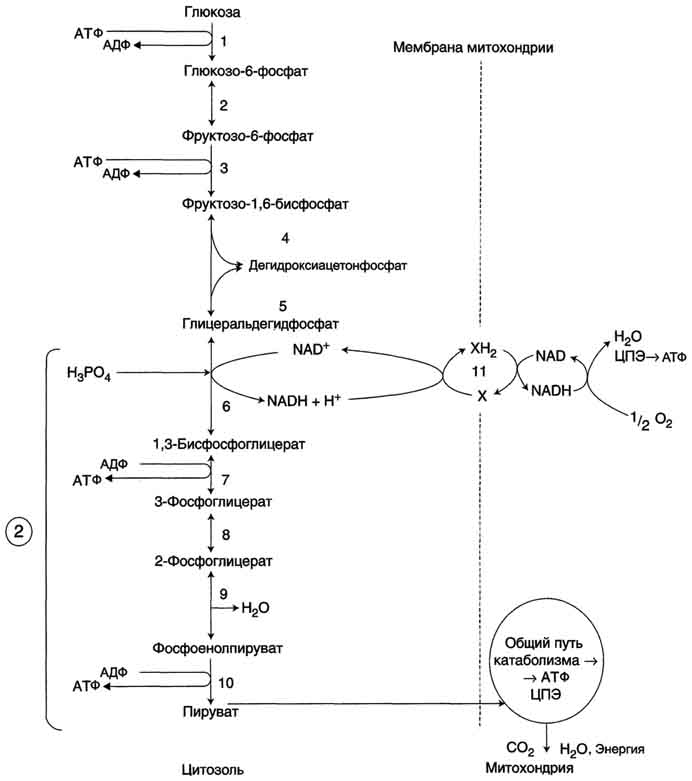
Аэробный распад глюкозы.
1-10- реакции аэробного гликолиза;
11 - малат-аспартатный челночный механизм транспорта водорода в митохондрии;
2 (В кружке) - стехиометрический коэффициент.
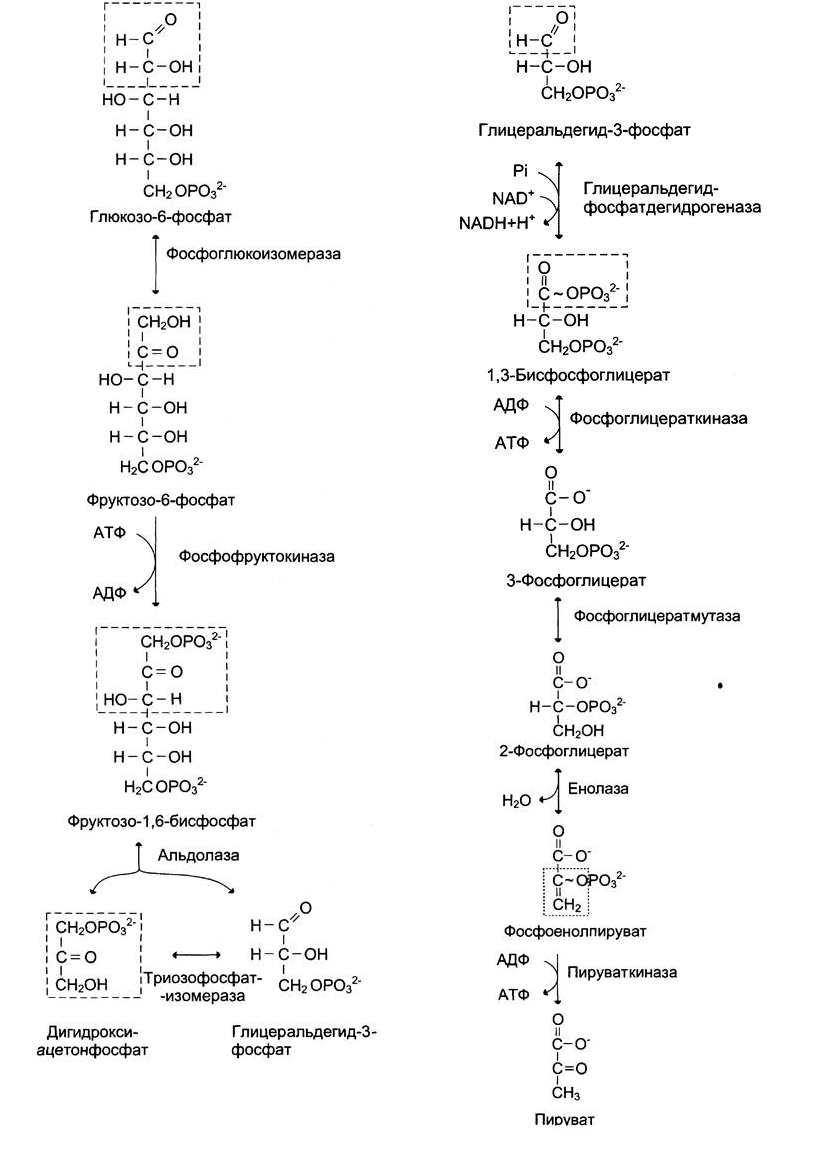
Реакции аэробного гликолиза:
1. Превращение глюкозо-6-фосфата превращается в фруктозо-6-фосфат
Фермент: глюкозофосфатизомераза
2. Фруктозо-6-фосфат превращается в фруктозо-1,6- бисфосфат.
Фермент: фосфофруктокиназа. Это наиболее медленная из всех реакций гликолиза, определяет скорость всего гликолиза, поэтому, регулируя активность фосфофруктокиназы, можно изменять скорость катаболизма глюкозы.
3-5. Фруктозо-1,6-бисфосфат далее расщепляется на 2 триозофосфата:
глицеральдегид-3-фосфат и дигидроксиацетонфосфат.
Фермент: фруктозобисфосфатальдолаза (или просто альдолаз). Этот фермент катализирует как реакцию альдольного расщепления, так и альдольной конденсации, т. е. обратимую реакцию. Продукты реакции альдольного расщепления – изомеры.
В последующих реакциях гликолиза используется только глицеральдегид-3-
фосфат, поэтому дигидроксиацетонфосфат под действием триозофосфатизомеразы превращается в глицероальдегид-3-фосфат
6. Превращение глицеральдегид-3-фосфата в 1,3- бисфосфоглицерат.
Это превращение – первая реакция окисления в ходе гликолиза.
Фермент: глицеральдегцд-3-фосфатдегидрогеназа (NAD-зависимая)
Окисление глицеральдегид-3-фосфата приводит к восстановлению NAD (нужен для
дыхательной цепи) и образованию с участием Н3РО4 высокоэнергетической ангидридной связи в 1,3-бисфосфоглицерате в положении 1.
7. Превращение 1,3-бисфосфоглицерата в 3-фосфоглицерат
Фермент: фосфоглицераткиназа
Высокоэнергетический фосфат передаётся на АДФ с образованием АТФ.
8. Превращение 3-фосфоглицерата в 2-фосфоглицерат
Фермент: фосфоглицератмутаза
Внутримолекулярные преобразования – перенос фосфатного остатка из положения 3 в 2.
9. Отщепление от 2-фосфоглицерата воды – обр. фосфоенолпируват
Фермент: енолаза. Название дегидратирующего фермента дано по обратной реакции.
Образованный фосфоенолпируват – макроэргическое соединение
10. Превращение фосфоенолпирувата в пируват
Фермент: пируваткиназа. Фосфатная гр. переносится на АДФ
Превращение фосфоенолпирувата в пируват – необратимая реакция. Это вторая в ходе гликолиза реакция субстратного фосфорилирования. Образующаяся енольная форма пирувата затем неферментативно переходит в более термодинамически стабильную кетоформу.
Физиологическое значение аэробного гликолиза:
Основное назначение катаболизма глюкозы – использование энергии,
освобождающейся при этом процессе для синтеза АТФ
Аэробный гликолиз происходит во многих тканях и явл. основным источником
энергии для жизнедеятельности (особенно важно для мозга)
Метаболиты гликолиза используются для синтеза других соединений
Использование глюкозы для синтеза жиров: ацетил-КоА, образующийся из
пирувата, используется как субстрат при биосинтезе жирных кислот, холестерина, а дигидроксиацетонфосфат как субстрат для синтеза глицерол-3-фосфата