Материал: Задачи по биохимии
45. Декарбоксилирование аминокислот. Образование и биологическая роль биогенных аминов: гамма-аминомасляной кислоты (гамк), гистамина, серотонина, дофамина. Инактивация биогенных аминов.
Внутримолекулярное дезаминирование
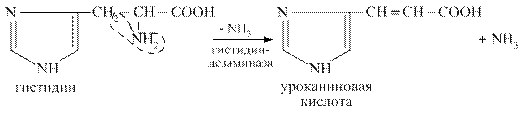
В процесс внутримолекулярного дезаминирования вступают аминокислоты гистидин, серин, треонин, цистеин. Например, из гистидина происходит выделение NН3 за счёт внутримолекулярной перестройки с образованием уроканиновой кислоты:
Серин в результате внутримолекулярного дезаминирования переходит в пировиноградную кислоту
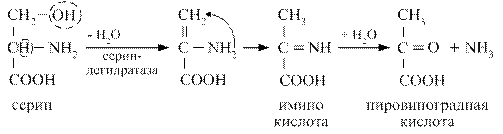
У детей процессы трансаминирования и дезаминирования идут более активно, чем у взрослых.
Декарбоксилирование аминокислот – ферментативный процесс высвобождения СО2 из СООН - групп аминокислот с образованием аминов.
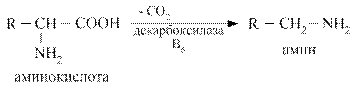
Наиболее активно в процесс деркарбоксилирования включаются аминокислоты гистидин, тирозин, глютамат, триптофан. Образующиеся амины называются биогенными аминами, поскольку они, как правило, обладают широким спектром физиологических эффектов, влияют на тонус сосудов, являются нейромедиаторами, участвуют в воспалительных реакциях. К основным биогенным аминам относятся гистамин, серотонин, катехоламины, гамма - аминомасляная кислота, полиамины.
Гистамин образуется при декарбоксилировании аминокислоты гистидина. Он синтезируется в тучных клетках, накапливается в секреторных гранулах, выделяется при раздражении клеток.
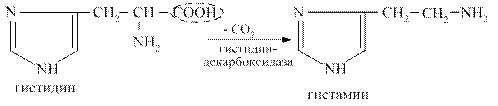
Гистамин оказывает разнообразные биологические эффекты: вызывает расширение сосудов, снижает артериальное давление, увеличивает тканевую проницаемость, вызывает местный отёк, стимулирует желудочную секрецию, обладает бронхоспатическим эффектом. В высокой концентрации он является медиатором воспалительных и аллергических реакций.
Серотонин образуется при декарбоксилировании гидрокситриптофана. Он синтезируется в хромаффиннных клетках кишечника, в некоторых ядрах подкорковых структур, в тромбоцитах.
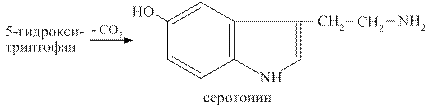
Эффекты серотонина: вызывает спазм сосудов, повышение артериального давления, стимулирует перистальтику кишечника, участвует в терморегуляции, в механизмах сна, памяти, является источником для синтеза гормона мелатонина, влияет на эмоциональные реакции человека.
Катехоламины (дофамин, адреналин, норадреналин) синтезируются из аминокислоты тирозина.
![]()
Дофамин – возбуждающий медиатор, при его дефиците развивается болезнь Паркинсона (адинамия, ригидность, тремор). Адреналин вызывает спазм сосудов, повышают артериальное давление, стимулирует работу сердца, является гормоном.
Норадреналин в основном выполняет нейромедиаторные функции.
Гамма - аминомасляная кислота (ГАМК) образуется при декарбоксилировании глютаминовой кислоты, является тормозным медиатором, улучшает кровоснабжение головного мозга, активирует окислительные процессы в нём.
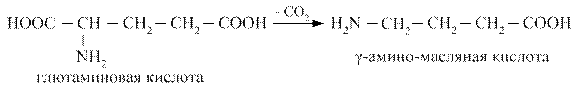
Полиамины (спермин, спермидин) синтезируются из орнитина и метионина, участвуют в регуляции процессов трансляции, транскрипции, репликации.
Так как биогенные амины очень активны, они быстро инактивируются в тканях. Распад биогенных аминов осуществляется несколькими способами: окисление, метилирование, дезаминирование. Основным способом инактивации биогенных аминов является окислительное дезаминирование под действием ферментов аминооксидаз (моноаминооксидаз, полиаминооксидаз).

Ингибиторы МАО применяются в качестве терапевтических средств.
46. Использование безазотистых остатков аминокислот: понятие, биологическая роль и примерыгликогенных и кетогенных аминокислот.
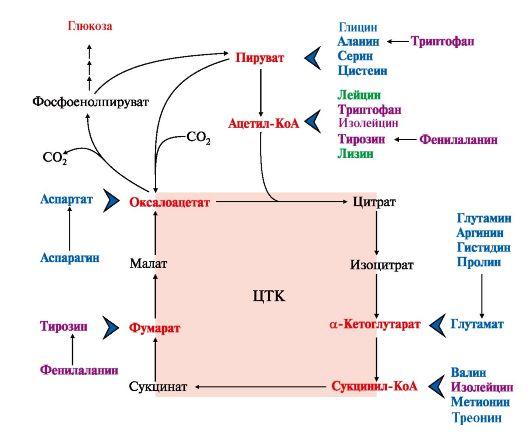
1. Катаболизм всех аминокислот сводится к образованию шести соединений, являющихся метаболитами ОПК (рис. 9.14):
• пируват,
• ацетил-КоА,
• α-кетоглутарат,
• сукцинил-КоА,
• фумарат,
• оксалоацетат.
Аминокислоты, которые в ходе катаболизма превращаются в пируват или другие промежуточные продукты ЦТК (α-КГ, сукцинил-КоА, фумарат), в итоге превращаются в оксалоацетат и используются в процессе глюконеогенеза. Эти аминокислоты образуют группу гликогенных аминокислот.
Образование глюкозы из аминокислот стимулирует гормон кортизол, который индуцирует в печени синтез ферментов глюконеогенеза, орнитинового цикла и АЛТ.
Некоторые аминокислоты в процессе катаболизма превращаются в ацетоацетат или ацетил-КоА и могут быть источником кетоновых тел - кетогенные аминокислоты: Лиз, Лей.
Катаболизм ряда аминокислот приводит к образованию соединений, которые могут использоваться для синтеза глюкозы, и кетоновых тел, так как превращаются сразу в два продукта - один из метаболитов ОПК и ацетилКоА (Иле) или ацетоацетат(Три, Фен, Тир). Такие аминокислоты называют смешанными или глико-кетогенными (см. рис. 9.14).
|
|
2. Полное окисление безазотистых остатков аминокислот до углекислого газа и воды реального энергетического значения не имеет. Основной путь их использования - включение в глюконеогенез. Этот процесс усиливается при голодании и сахарном диабете.
Безазотистые остатки аминокислот используются для восполнения количества метаболитов ОПК, которое затрачивается на синтез биологически активных веществ
Такие реакции называют анаплеротическими. Можно выделить пять анаплеротических реакций:
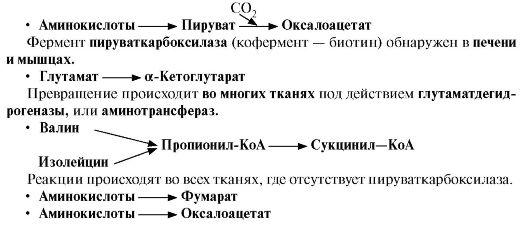 Последние две
реакции происходят в печени.
Последние две
реакции происходят в печени.
Первые две реакции являются основными анаплеротическими реакциями
47. Источники образования аммиака в организме. Причины токсичности аммиака. Понятие гипераммониемии.
1. Основным источником аммиака является катаболизм аминокислот в
тканях. Небольшая часть аммиака образуется в клетках при распаде азотсодержащих соединений (биогенных аминов, нуклеотидов и др.) (рис. 9.7), а также при гниении белков в кишечнике в результате деятельности микрофлоры, откуда он частично всасывается и поступает в воротную вену. Концентрация аммиака в крови воротной вены существенно выше, чем в общем кровотоке.
|
|
Катаболизм аминокислот и образование аммиака происходит во всех тканях организма. Однако концентрация аммиака в крови очень мала, так как он быстро связывается в клетках с образованием нетоксичных продуктов. Содержание аммиака в крови в норме составляет всего 0,4-0,7 мг/л (25-40 мкмоль/л).
Из организма аммиак выводится почками в виде конечных продуктов азотистого обмена:
• мочевины -
синтезируется в печени;• аммонийных
солей -
образуются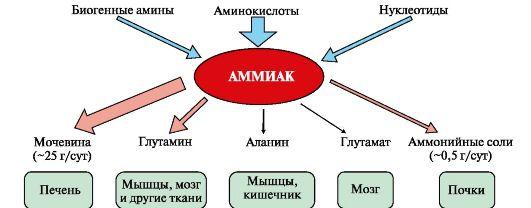 в почках.
в почках.
Причины токсичности аммиака
1. Аммиак сдвигает реакцию, катализируемую глутаматдегидрогеназой, в сторону образования глутамата. В результате снижается концентрация -кетоглутарата, что вызывает угнетение обмена аминокислот (переаминирова-ние) и гипоэнергетическое состояние (угнетение цикла Кребса).
2. Аммиак усиливает синтез глутамина из глутамата в нервной ткани. Снижение концентрации глутамата нарушает обмен нейромедиаторов, в частности синтез -аминомасляной кислоты – основного тормозного модуля-тора. Это нарушает проведение нервного импульса и вызывает судороги.
3. Аммиак в крови и цитозоле образует ион NH4+, накопление которого нарушает трансмембранный перенос ионов, в частности Na+ и К+, что также влияет на проведение нервного импульса.
4. Накопление аммиака может сдвигать рН в щелочную сторону, вызывая метаболический алкалоз.
Гипераммониемия - это наследственное заболевание обмена веществ, проявляющееся в недостаточности цикла ферментов мочевины, приводящее к отравлению организма аммиаком. Аммиак является токсичным соединением, находящимся в крови в относительно небольших концентрациях (11,0-32,0 мкмоль/л). Симптомы аммиачного отравления проявляются при превышении этих пределов всего в 2-3 раза. Предельно допустимый уровень аммиака в крови 60 мкмоль/л. При повышении концентрации аммиака (гипераммониемия) до предельных величин может наступить кома и смерть. При хронической гипераммониемии развивается умственная отсталость.
48. Пути обезвреживания аммиака и их особенности в почках, печени, нервной ткани.
В
тканях аммиак находится преимущественно
в виде иона аммония NН![]() в
равновесии с небольшой концентрацией
неионизированного аммиака.
в
равновесии с небольшой концентрацией
неионизированного аммиака.
1. Восстановительное аминирование.
Одним из путей связывания и обезвреживания аммиака в организме, в частности в мозге, сетчатке, почках, печени и мышцах - это биосинтез амидов глутаминовой и аспарагиновой кислот (глутамина или аспарагина):
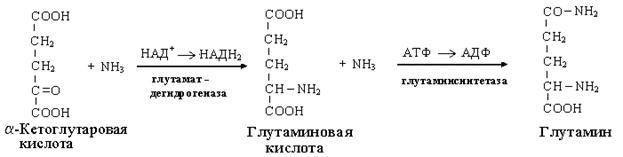
Эта реакция протекает во многих тканях, но наиболее важна для нервной, особенно чувствительной к токсическому действию аммиака. Первая реакция представляет собой обращение глутаматдегидрогеназной реакции (обратная окислительному дезаминированию ГЛУ).
Обезвреживание аммиака путем синтеза глутамина имеет и анаболическое значение, поскольку глутамин используется для синтеза ряда соединений. Прежде всего нужно отметить, что глутамин — одна из 20 аминокислот, входящих в белки. Кроме того, амидная группа глутамина используется для синтеза аспарагина, глюкозамина и других аминосахаров, пуриновых и пиримидиновых нуклеотидов. Таким образом, в этих реакциях азот аммиака включается в разнообразные структурно-функциональные компоненты клетки.
Глутамин затем может поступать во все ткани, где осуществляется его гидролиз при участии глутаминазы:
![]()
Подобным образом происходит образование аспарагина (через ЩУК).
2. Образование аммонийных солей.
Экскреция аммиака с мочой в норме невелика — около 0,5 г в сутки. Но она в несколько раз повышается при ацидозе, т. е. при увеличении содержания кислот в организме. Аммиак в почках образуется главным образом за счет амидной группы глутамина. Глутамин гидролизуется активируемой фосфатом глутаминазой, имеющейся в клетках эпителия канальцев почки. Часть аммиака (примерно 30%) образуется другим путем — в результате непрямого дезаминирования аминокислот.
Образующийся аммиак нейтрализует кислоты: NH3 + Н+ → NH4+. Неионизированные аммиак и кислоты в клетках находятся в равновесии с их ионизированными формами. Через клеточные мембраны проникают преимущественно неионизированные аммиак и кислоты, и в просвете почечного канальца (т. е. уже в моче) аммиак акцептирует протон кислоты, образуя аммонийную соль, которая выводится из организма. Экскреция аммиака почками служит для выведения именно кислот, а не азота, на что указывает значительная скорость экскреции при ацидозе, малая — при нормальной кислотности межклеточной жидкости и крови, и отсутствие экскреции аммиака при алкалозе. Одновременно этот процесс обеспечивает сохранение организмом ионов Na+, которые в отсутствие ионов аммония выводились бы с анионами кислот. Потеря таких количеств Na+, которые необходимы для выведения кислот при ацидозе, могла бы вызвать снижение осмотического давления межклеточной жидкости и крови, а вследствие этого уменьшение объема межклеточной жидкости, т. е. обезвоживание тканей.
3. Основным механизмом связывания аммиака в организме является синтез мочевины. Мочевина выводится из организма с мочой в качестве главного конечного продукта белкового, соответственно аминокислотного обмена. На долю мочевины приходится до 80-85 % от всего выводимого из организма азота. Количество выделяемой мочевины зависит от количества белков, поступающих с пищей. Если суточный рацион включает 80–100 г белка, то за сутки образуется и выводится 25–30 г мочевины.
Основным местом синтеза мочевины является печень. Синтез мочевины является циклическим метаболическим процессом и носит название орнитинового цикла мочевинообразования Кребса (цикл мочевины Кребса - Хензеляйта).
На первом этапе из NН3 и СО2 при участии АТФ синтезируется макроэргическое соединение карбамоилфосфат: