Материал: Задачи по биохимии
37. Синтез и использование кетоновых тел. Понятие и причины кетонемии и кетонурии.
Под термином «кетоновые (ацетоновые) тела» подразумевают ацетоук-суснуюкислоту (ацетоацетат) СН3СОСН2СООН, β-оксимасляную кислоту (β-оксибутират, или D-3-гидроксибутират) СН3СНОНСН2СООН и ацетон СН3СОСН3.
В здоровом организме ацетон в крови присутствует в крайне низких концентрациях, образуется в результате спонтанного декарбоксилирования ацетоацетата и, по-видимому, не имеет определенного физиологического значения.
Кетоновые тела образуются в печени. Прежние представления о том, что кетоновые тела являются промежуточными продуктами β-окисления жирных кислот, оказались ошибочными.
Во-первых, в обычных условиях промежуточными продуктами β-окис-ления жирных кислот являются КоА-эфиры этихкислот, например β-окси-бутирил-КоА, ацетоацетил-КоА.
Во-вторых, β-оксибутирил-КоА, образующийся в печени при β-окисле-нии жирных кислот, имеет L-конфигурацию, в то время как β-оксибутират, обнаруживаемый в крови, представляет собой D-изомер. Именно β-окси-бутират D-конфигурации образуется в ходе метаболического пути синтеза β-окси-β-метилглутарил-КоА (3-гидрокси-3-метилглутарил-КоА):
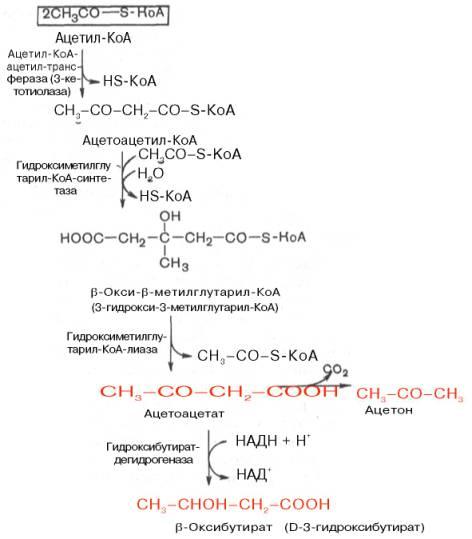
На первом этапе из 2 молекул ацетил-КоА образуется ацетоацетил-КоА. Реакция катализируется ферментом ацетил-КоА-ацетилтрансферазой (3-ке-тотиолазой). Затем ацетоацетил-КоА взаимодействует еще с одной молекулой ацетил-КоА. Реакция протекает под влиянием фермента гидро-ксиметилглутарил-КоА-синтетазы. Образовавшийся β-окси-β-метилглута-рил-КоА способен под действием гидроксиметилглутарил-КоА-лиазы расщепляться на ацетоацетат и ацетил-КоА.
Ацетоацетат восстанавливается при участии НАД-зависимой D-3-гид-роксибутиратдегидрогеназы, при этом образуется D-β-оксимасляная кислота (D-3-гидроксибутират). Следует подчеркнуть, что фермент специфичен по отношению к D-стереоизомеру и не действует на КоА-эфиры.
Существует второй путь синтеза кетоновых тел. Образовавшийся путем конденсации 2 молекул ацетил-КоА ацетоацетил-КоА способен отщеплять коэнзим А и превращаться в ацетоацетат. Этот процесс катализируетсяферментом ацетоацетил-КоА-гидролазой (деацилазой):
![]()
Однако второй путь образования ацетоуксусной кислоты (ацетоацетата) не имеет существенного значения, так какактивность деацилазы в печени низкая.
В настоящее время ясна молекулярная основа изречения, что «жиры сгорают в пламени углеводов». Известно, что ацетил-КоА, образовавшийся при окислении жирных кислот, включается в цикл трикарбоновых кислот в условиях, когда расщепление жиров и углеводов соответствующим образом сбалансировано. Включение ацетил-КоА в цикл Кребса зависит от доступности оксалоацетата для образования цитрата. Однако если расщепление жировпреобладает, судьба ацетил-КоА изменяется. Объясняется это тем, что в отсутствие углеводов или при нарушении их использования концентрация оксалоацетата снижается. При голодании или диабете окса-лоацетат расходуется на образование глюкозы и поэтому не может конденсироваться с ацетил-КоА. В таких условиях путь метаболизма аце-тил-КоА отклоняется в сторону образования ацетоацетата и D-3-гидрокси-бутирата, т.е. кетоновых тел.
В крови здорового человека кетоновые тела содержатся лишь в очень небольших концентрациях (в сыворотке крови0,03–0,2 ммоль/л). При патологических состояниях (у лиц с тяжелой формой сахарного диабета, при голодании, а также у животных с экспериментальным острым стрептозотоциновым или аллоксановым диабетом) концентрациякетоновых тел в сыворотке крови увеличивается и может достигать 16–20 ммоль/л.
Следует подчеркнуть важную роль кетоновых тел в поддержании энергетического баланса. Кетоновые тела – поставщики «топлива» для мышц, почек и действуют, возможно, как часть регуляторного механизма с обратной связью, предотвращая чрезвычайную мобилизацию жирных кислот из жировых депо. Печень в этом смысле является исключением, она не использует кетоновые тела в качестве энергетического материала.
38. Липопротеины сыворотки крови: биологическая роль, особенности состава и разновидности. Понятие атерогенных и антиатерогенных липопротеинов.
Липиды не растворимы в воде и транспортируются в крови в составе липопротеинов
В сферических частицах липопротеинов неполярные липиды — эфиры холестерина и триглицериды образуют гидрофобное ядро, полярные липиды — холестерин и фосфолипиды совместно с амфипатическими белками — аполипопротеинами располагаются в гидрофильном поверхностном слое. Аполипопротеины придают липопротеинам стабильность, выполняют функцию лигандов клеточных рецепторов, определяют метаболическую судьбу липопротеинов.
Липопротеины участвуют в транспорте липидов пищи (экзогенный путь) и липидов, синтезированных в организме (эндогенный путь).
Экзогенный путь — всасывание липидов пищи в тонком кишечнике с помощью хиломикронов. Они образуются в энтероцитах, состоят из пищевых триглицеридов в комплексе с фосфолипидами и аполипопротеином В-48, поступают в кровоток через лимфатический грудной проток. После секреции хиломикроны получают аполипопротеины А-I, A-II, A-IV, С-II/С-III и Е. В крови триглицериды хиломикронов подвергаются липолизу под влиянием фермента эндотелия сосудов — липопротеидлипазы. Хиломикроны с истощенным содержанием триглицеридов становятся остатками хиломикронов (ремнанты). Эти остатки захватываются гепатоцитами при участии рецепторов апо В/Е.
Эндогенная система включает липопротеины очень низкой плотности (ЛПОНП), липопротеины промежуточной плотности, липопротеины низкой плотности (ЛПНП) и липопротеины высокой плотности (ЛПВП).
ЛПОНП синтезируются в печени. Они обогащены триглицеридами, содержат холестерин и аполипопротеины. Для секреции ЛПОНП из гепатоцитов в кровь необходим апоВ-100, затем в крови присоединяются апо Е и апо С-II/С-III. Триглицериды ЛПОНП гидролизуются эндотелиальной липопротеидлипазой (ее активатор — апо С-II). Жирные кислоты триглицеридов используются для ресинтеза жира в жировой ткани или окисляются в скелетных мышцах. Часть ЛПОНП превращается в ЛПНП через стадию липопротеинов промежуточной плотности.
Таблица 74.Классы липопротеинов
|
Класс липопротеинов |
Плотность, г/мл |
Диаметр, нм |
Основные липиды |
Основные аполипо-протеины |
Подвижность при электрофорезе |
|
Хиломикроны |
<0,95 |
80 — 120 |
Пищевые триглицериды |
Апо B-48 Апо А-I Апо A-II Апо A-IV Апо C-II/C-III Апо E |
Остаются на линии старта |
|
ЛПОНП |
< 1,006 |
30 — 80 |
Эндогенные триглицериды |
Апо В-100 Апо E Апо C-II/C-III |
Пре-β |
|
Липопротеины промежуточной плотности |
1,006 — 1,019 |
23 — 35 |
Эфиры холестерина, триглицериды |
Апо В-100 Апо E Апо C-II/C-III |
Медленные пре-β |
|
ЛПНП |
1,019 — 1,063 |
16 — 25 |
Эфиры холестерина |
Апо В-100 |
β |
|
ЛПВП |
1,063 — 1,210 |
5 — 12 |
Эфиры холестерина, фосфолипиды |
Апо А-I Апо A-II Апо C-II/C-III |
a |
|
Липопротеин (а) |
1,055 — 1,086 |
|
Эфиры холестерина |
Апо В-100 Апо (а) |
Медленный пре-β |
Как отмечалось, основным местом образования ацетоацетата и 3-гид-роксибутирата служит печень. Из митохондрийпечени эти соединения диффундируют в кровь и переносятся к периферическим тканям.
Действительно, сердечная мышца и корковый слой почек предпочтительно используют в качестве «топлива» ацетоацетат, а не глюкозу.
В противоположность этому глюкоза является главным «топливом» для мозга у лиц, получающих сбалансированную пищу. При голодании и диабете мозг адаптируется к использованию ацетоацетата. Установлено, что в условиях длительного голодания 75% потребности мозга в «топливе» удовлетворяется за счет ацетоацетата.
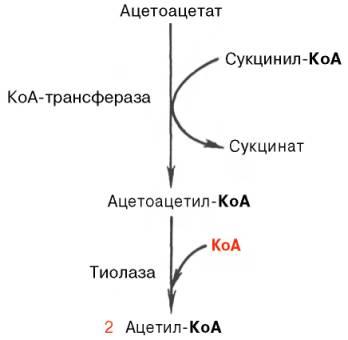
Известно, что в периферических тканях 3-гидроксибутират (β-оксимас-ляная кислота) способен окисляться до ацетоацетата, а последний активируется с образованием соответствующего КоА-эфира (ацетоацетил-КоА). Ацетоацетат может быть активирован путем переноса КоА с сукцинил-КоА в реакции, катализируемой специфической КоА-трансферазой. Образовавшийся ацетоацетил-КоА далее расщепляется тиолазой с образованием 2 молекулацетил-КоА, которые затем включаются в цикл Кребса:
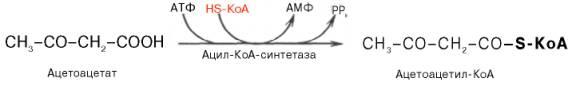
Не исключено, что существует и второй путь активации ацетоацетата – это использование АТФ и HS-KoA аналогично тому, как при активации жирных кислот:
Примечание. ЛПОНП — липопротеины очень низкой плотности, ЛПНП — липопротеины низкой плотности, ЛПВП — липопротеины высокой плотности.