Материал: В-Л-Быков-Цитогогия_и_общая_гистология
Форма эпителиоцитов служит важным классификационным признаком как отдельных клеток, так и эпителиальных пластов в целом. Выделяют плоские, кубические и призматические (столбчатые, или цилиндрические) клетки. Эпителиоцитам, как уже отмечено выше, свойственна полярность.
Ядро зпителиоцитов может иметь различную форму, которая обычно соответствует форме клетки: в плоских клетках оно дисковидное, в кубических - сферическое, в цилиндрических - эллипсоидное. В большинстве клеток ядро сравнительно светлое (преобладает эухроматин), содержит хорошо заметное крупное ядрышко, однако в ороговевающих эпителиях по мере дифференцировки клеток оно уменьшается, уплотняется, распадается и лизируется - подвергается кариопикнозу, кариорексису и кариолизису (термины см. главу 3).
Цитоплазма зпителиоцитов содержит все органеллы общего значения, а в некоторых клетках - также органеллы специального значения, обеспечивающие функции данных клеток. В клетках железистого эпителия хорошо развит синтетический аппарат. В связи с полярностью клеток органеллы распределены в их цитоплазме неравномерно.
Цитоскелет зпителиоцитов хорошо развит и представлен микротрубочками, микрофиламентами и промежуточными филаментами. Последние в эпителиоцитах особенно многочисленны и называются тонофиламентами. При фиксации, склеиваясь друг с другом, они могут образовывать крупные агрегаты, выявляемые под световым микроскопом и описанные под названием тонофибрилл.
Цитокератины - белки, образующие тонофиламенты, которые специфичны для клеток эпителиальных тканей. Исключение составляет эндотелий, для которого характерны виментиновые промежуточные филаменты. Идентифицировано около 30 различных форм цитокератинов. Которые представлены двумя типами: кислыми кератинами (тип I) и основными кератинами (тип II). Выработка каждого вида цитокератина кодируется особым геном. Тонофиламенты образованы гетерополимерным комплексом, включающим не менее двух различных цитокератинов (кислый + основной). Для конкретного вида эпителия (а в многослойных эпителиях - для каждого слоя) характерен определенный набор цитокератинов, экспрессию которых рассматривают как маркер дифференцировки эпителиальных клеток.
Изменения нормальной экспрессии цитокератинов могут указывать на нарушения дифференцировки клеток и в ряде случаев служить
- 121 -
важным диагностическим признаком их злокачественного перерождения. При ряде заболеваний кожи и слизистых оболочек в эпитслиопитах обнаружены изменения кератинов, связанные с мутациями соответствующих генов.
Поверхности эпителиоцита (латеральная, базальная, апикальная)
обладают отчетливой структурно-функциональной специализацией, которая особенно хорошо выявляется в однослойном эпителии.
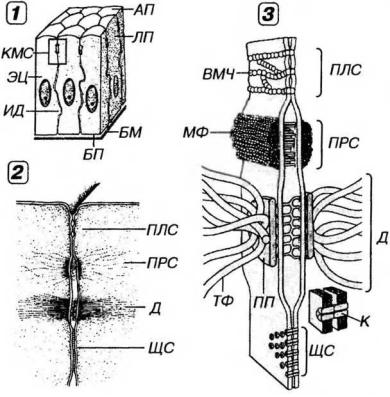
Рис. 5-2. Межклеточные соединения эпителиоцитов в области их латеральной поверхности. 1 - область расположения комплекса межклеточных соединений (выделена рамкой), 2 - вид межклеточных соединений на ультратонких срезах, 3 - трехмерная схема строения межклеточных соединений (по К.Де Дюву, 1987, с изменениями). БМ - базальная мембрана, БП - базальная поверхность, АП - апикальная поверхность, ЛП - латеральная поверхность эпителиоцитов, КМС - комплекс межклеточных соединений, ПЛС - плотное соединение, ПРС - промежуточное соединение, Д - десмосома, ИД - интердигитации, ВМЧ - внутримембранные частицы, ПП - пластинка прикрепления, МФ - микрофиламенты. ТФ - тонофиламенты, ЩС - щелевое соединение, К - коннексоны.
- 122 -
Латеральная поверхность зпителиоцитов обеспечивает связь клеток друг с другом за счет специализированных участков - межкле точных соединений, или контактов (рис. 5-2). Благодаря последних эпителиоциты формируют пласты, что служит важнейшим отличительным свойством организации эпителиальных тканей.
Межклеточные соединения подразделяются на два основных вида:
1.Механические соединения - обусловливают механическую связь зпителиоцитов друг с другом. В их число входят плотные соединения, промежуточные соединения, десмосомы, интердигитации;
2.Коммуникационные соединения - (от лат. communicatio - сообщение)
обеспечивают химическую (метаболическую, ионную и электрическую) связь между эпителиоцитами. К ним относятся щелевые соединения.
(1) Плотное соединение (zonula occludens - поясок замыкания) -
наиболее тесный контакт клеток из всех известных в природе. Представляет собой область частичного слияния наружных листков плазмолемм двух соседних клеток (см. рис. 5-2), которая блокирует распространение веществ по межклеточному пространству (обеспечивая тем самым барьерную функцию эпителия и регулируемость транспорта веществ через эпителиальный пласт). Это соединение также препятствует свободному перемещению и смешиванию функционально различных внутримембранных белков, локализующихся в плазмолемме апикальной и базолатеральной поверхностей клетки, что способствует поддержанию ее полярности.
Плотное соединение имеет вид пояска шириной 0.1-0.5 мкм, окружающего клетку по периметру (обычно у ее апикального полюса) и состоящего из анастомозирующих тяжей внутримембранных частиц. Эти частицы образованы белком окклюдином, каждая из них представляет собой область точечного слияния плазмолемм двух соседних клеток. Проницаемость плотных соединений тем ниже, чем выше число тяжей таких частиц. Для поддержания целостности этих соединений необходимы двухвалентные катионы (Са2+, Mg2+). Они метут динамично перестраиваться (вследствие изменений экспрессии и степени полимеризации окклюдина) и временно размыкаться (например, при миграции лейкоцитов через межклеточные пространства).
(2) Промежуточное соединение, или опоясывающая десмосома (zonula adherens - поясок сцепления) локализуется на латеральной
- 123 -
поверхности эпнтелиоцнта между областью расположения плотного соединения и десмосом (что обусловило его первое название). Охватывает клетку по периметру в виде пояска (см. рис. 5-2), на сечении имеющего сходство с десмосомой (что послужило основанием для второго названия). В области промежуточного соединения обращенные к цитоплазме листки плазмолеммы утолщены, образуя пластинки прикрепления, которые содержат актинсвязывающие белки α-актинин, винкулин и плакоглобин (последний обнаруживается также в десмосомах). К этим пластинкам прикрепляются элементы цитоскелета - актиновые микрофиламенты, вплетающиеся также в терминальную сеть. Межклеточная щель расширена до 15-20 нм и заполнена умеренно плотным веществом, в состав которого входит адгезивный трансмембранный гликопротеин Е-кадгерин, обеспечивающий в присутствии ионов Са2+ связь между соседними клетками. Со стороны цитоплазмы в области промежуточного соединения к Е-кадгерину через α -актинин и винкулин прикрепляются актиновые микрофиламенты, что обусловливает связь цитоскелета с компонентами межклеточного вещества.
(3) Десмосома (macula adherens - пятно сцепления) - состоит из утолщенных и уплотненных участков цитоплазматического листка плазмолемм двух соседних клеток - пластинок прикрепления, разделе-ных межклеточной щелью (см.рис. 5-2).
Пластинки прикрепления имеют дисковидную форму (диаметр около 0.5 мкм, толщина 15 нм) и служат участками прикрепления к плазмолемме промежуточных филаментов (тонофиламентов). Они содержат особые белки - десмоплакины, плакоглобин и десмокальмин.
Межклеточная щель в области десмосомы имеет ширину около 25 нм и заполнена материалом низкой электронной плотности, часто поперечно исчерченным и содержащим в центре линейное уплотнение (центральная, или промежуточная линия). В межклеточном материале десмосомы находятся десмоколлины и десмоглеины - трансмембранные Са2+-связывающие адгезивные белки, которые, взаимодействуя с белками пластинок прикрепления, связывают их в единую систему.
Десмосомы разбросаны по поверхности клетки; они, как и промежуточные соединения, служат участками, опосредующими связь элементов цитоскелета (внутриклеточного компонента) с компонентами межклеточного вещества.
Повреждение десмосом посредством антител к их компонентам служит главным механизмом патогенеза вульгарной пузырчатки - тяжелого, а в прежние годы смертельного аутоиммунного заболевания кожи и слизистых оболочек. Клетки в многослойных эпителиях утрачивают связи друг с другом, округляются, а их тонофиламенты отсоединяются
- 124 -
от пластинок прикрепления и образуют скопления вокруг ядра. Внутри эпителия формируются пузыри, которые вскрываются с образованием эрозий. Связь эпителия с подлежащей соединительной тканью, однако, при этом заболевании сохраняется.
(4)Интердигитации - межклеточные соединения, образованные выпячиваниями цитоплазмы одних клеток, вдающимися в цитоплазму других (см. рис. 5-2). За счет интердигитаций увеличивается прочность соединения клеток друг с другом и нарастает площадь поверхности, через которую могут осуществляться межклеточные обменные процессы.
(5)Щелевое соединение (nexus) образовано совокупностью трубчатых трансмембранных структур диаметром 9-11 нм (коннексонов), пронизывающих плазмолеммы соседних клеток на участках диаметром 0.5-3 мкм и стыкующихся друг с другом в области узкой межклеточной щели шириной 2-3 нм (см. рис. 5-2). Число коннексонов в щелевом соединении обычно исчисляется сотнями. Каждый коннексон представлен 6 (иногда 4 или 5) субъединицами, образованными белком коннексином, и пронизан каналом диаметром 1.5-2.0 нм, который обусловливает свободный обмен низкомолекулярными (с массой до 2 кД) соединениями (неорганическими ионами, сахарами, витаминами, аминокислотами, нуклеотидами, АТФ и др.) между клетками, обеспечивая их ионное и метаболическое сопряжение.
Базальная поверхность эпителиоцитов прилежит к базальной мембране, к которой она прикреплена с помощью полудесмосом - соединений, сходных по строению с половинами десмосом. Молекулярная организация и биохимический состав полудесмосом и десмосом, однако, не идентичны. В функциональном плане базальная и латеральная (до уровня плотных соединений) части плазмолеммы эпителиоцита в совокупности образуют единый комплекс (базо-латеральную плазмолемму), который содержит специфические для него мембранные белки. Эти белки служат (а) рецепторами, воспринимающими различные сигнальные молекулы (гормоны, факторы роста), (б) переносчиками питательных веществ, поступающих из сосудов подлежащей соединительной ткани, (в) ионными насосами и др.
Базальная поверхность может быть сравнительно плоской или образовывать выроста - базальные отростки. Последние могут служить для обеспечения более прочной связи эпителиоцитов с соединительной тканью путем увеличения площади поверхности их соприкосновения (например, в многослойных эпителиях).
- 125 -