Материал: В-Л-Быков-Цитогогия_и_общая_гистология
Расположение микротрубочек. Микротрубочки располагаются в цитоплазме в составе нескольких систем:
а) в виде отдельных элементов, разбросанных по всей цитоплазме и формирующих сети;
б) в пучках, где они связаны тонкими поперечными мостиками (в отростках нейронов, в составе митотического веретена, манжетки сперматиды, периферического "кольца" тромбоцитов);
в) частично сливаясь друг с другом с формированием пар, или дyблетов (в аксонеме ресничек и жгутиков), и триплетов (в базальном тельце и центриоли).
Образование и разрушение микротрубочек. Микротрубочки представляют собой лабильную систему, в которой имеется равновесие между их постоянной сборкой и диссоциацией. У большинства микротрубочек один конец (обозначаемый как "-") закреплен, а другой ("+") свободен и участвует в их удлинении или деполимеризации. Структурами, обеспечивающими образование микротрубочек, служат особые мелкие сферические тельца - сателлиты (от англ. satellite - спутник), отчего последние называют центрами организации микротрубочек (ЦОМТ). Сателлиты содержатся в базальных тельцах ресничек и клеточном центре (см. рис. 3-15 и 3-16). После полного разрушения микротрубочек в цитоплазме они отрастают от клеточного центра со скоростью около 1 мкм/мин., а их сеть вновь восстанавливается менее, чем за полтора часа. К ЦОМТ относят также и центромеры хромосом.
Связь микротрубочек с другими структурами клетки и между собой
осуществляется посредством ряда белков, выполняющих различные функции.
(1) Микротрубочки с помощью вспомогательных белков прикреплены к другим клеточным компонентам. (2) По своей длине микротрубочки образуют многочисленные боковые выросты (которые состоят из белков, ассоциированных с микротрубочками) длиной до нескольких десятков нанометров. Благодаря тому, что такие белки последовательно и обратимо связываются с органеллами, транспортными пузырьками, секреторными гранулами и другими образованиями, микротрубочки (которые сами не обладают сократимостью) обеспечивают перемещение указанных структур по цитоплазме. (3) Некоторые белки, ассоциированные с микротрубочками, стабилизируют их структуру, а связываясь с их свободными краями, препятствуют деполимеризации.
Угнетение самосборки микротрубочек посредством ряда веществ,
являющихся ингибиторами митоза (колхицин, винбластин, винкристин), вызывает избирательную гибель быстроделящихся клеток. Поэтому некоторые из таких веществ успешно используются для химиотера-
- 66 -
паи опухолей. Блокаторы микротрубочек нарушают также транспортные процессы в цитоплазме, в частности, секрецию, аксонный транспорт в нейронах. Разрушение микротрубочек приводит к изменениям формы клетки и дезорганизации ее структуры и распределения органелл.
Клеточный центр (цитоцентр)
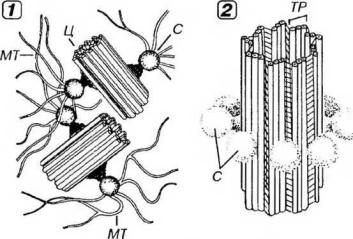
Клеточный центр образован двумя полыми цилиндрическими структурами длиной 0.3-0.5 мкм и диаметром 0.15-0.2 мкм - центриолями, которые располагайся вблизи друг друга во взаимно перпендикулярных плоскостях (рис. 3-15). Каждая центриоль состоит из 9 триплетов частично слившихся микротрубочек (А, В и С), связанных поперечными белковыми мостиками ("ручками"). В центральной часта центриоли микротрубочки отсутствуют (по некоторым данным, здесь имеется особая центральная нить), что описывается общей формулой (9*3) + 0. Каждый триплет центриоли связан со сферическими тельцами диаметром 75 нм - сателлитами-, расходящиеся от них микротрубочки образуют центросферу.
Рис. 3-15. Клеточный центр (1) и структура центриоли (2). Клеточный центр образован парой центриолей (Ц), расположенных во взаимноперпендикулярных плоскостях. Каждая Ц состоит из 9 связанных друг с другом триплетов (ТР) микротрубочек (МТ). С каждым ТР посредством ножек связаны сателлиты (С) - глобулярные белковые тельца, от которых отходят МТ.
В неделящейся клетке выявляется одна пара центриолей (диплосома), которая обычно располагается вблизи ядра. Перед делением в S-периоде интерфазы происходит дупликация центриолей пары, причем под
- 67 -
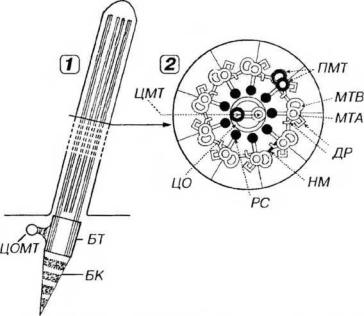
Рис. 3-16. Ресничка. 1 - продольный срез, 2 - поперечный срез. БТ - базальное тельце (образовано триадами микротрубочек), ЦОМТ - центр организации микротрубочек, БК - базальный корешок, ПЛ - плазмолемма, МТА - микротрубочка А, МТБ - микротрубочка В, ПМТ - периферические микротрубочки, ЦМТ - центральные микротрубочки, ЦО - центральная оболочка, ДР - динеиновые ручки, PC - радиальные спицы, НМ - нексиновые мостики.
прямым углом к каждой зрелой (материнской) центриоли формируется новая (дочерняя), незрелая процентриоль, в которой вначале имеются лишь 9 единичных микротрубочек, позднее превращающихся в триплеты. Пары ценгриолей далее расходятся к полюсам клетки, а во время митоза они служат центрами образования микротрубочек ахроматинового веретена деления.
Реснички и жгутики
Реснички и жгутики - органеллы специального значения, участвующие в процессах движения, - представляют собой выросты цитоплазмы, основу которых составляет каркас из микротрубочек, называемый осевой нитью, или аксонемой (от греч. axis - ось и пета - нить). Длина ресничек равна 2-10 мкм, а их количество на поверхности одной реснитчатой клетки может достигать нескольких сотен. В единственном типе клеток человека, имеющих жгутик - спермиях - содержится только по одному жгутику длиной 50-70 мкм.
- 68 -
Аксонема образована 9 периферическими нарами микротрубочек и одной центрально расположенной нарой; такое строение описывается формулой (9 х 2) + 2 (рис. 3-16). Внутри каждой периферической пары за счет частичного слияния микротрубочек одна из них (А) полная, а вторая (В) - неполная (2-3 димера общие с микротрубочкой А).
Центральная пара микротрубочек окружена центральной оболочкой, от которой к периферическим дублетам расходятся радиальные спицы. Периферические дублеты связаны друг с другом мостиками нексина, а от микротрубочки А к микротрубочке В соседнего дублета отходят "ручки" из белка динеина (см. рис. 3-16), который обладает активностью АТФазы.
Биение реснички и жгутика обусловлено скольжением соседних дублетов в аксонеме, которое опосредуется движением динеииовых ручек. Мутации, вызывающие изменения белков, входящих в состав ресничек и жгутиков, приводят к различным нарушениям функции соответствующих клеток. При синдроме Картагенера (синдроме неподвижных ресничек), обычно обусловленном отсутствием динеииовых ручек, больные сградаюг хроническими заболеваниями дыхательной системы (связанными с нарушением функции очищения поверхности респираторного эпителия) п бесплодием (вследствие неподвижности спермиев).
Базальное тельце, по своему строению сходное с цеитриолью, лежит в основании каждой реснички или жгутика. На уровне апикального конца тельца микротрубочка С триплета заканчивается, а микротрубочкн А и В продолжаются в соответствующие микротрубочки аксонемы реснички или жгутика. При развитии ресничек или жутка базальное тельце играет роль матрицы, на которой поисходит сборка компонентов аксонемы.
Микрофиламенты
Микрофиламенты - тонкие белковые нити диаметром 5-7 нм, лежащие в цитоплазме поодиночке, в виде сетей ими пучками. В скелетной мышце тонкие микрофиламенты образуют упорядоченные пучки, взаимодействуя с более толстыми миознновыми филаментами.
Кортикальная (терминальная) сеть - зона сгущения микрофиламентов под плазмолеммой, характерная для большинства клеток. В этой сети микрофиламенты переплетены между собой и "сшиты" друг с другом с помощью особых белков, самым распространенным из которых является филамин. Кортикальная сеть препятствует резкой и внезапной деформации клетки при механических воздействиях и обеспечивает плавные изменения ее формы путем перестройки, которая облегчается актин-растворяющими (преобразующими) ферментами.
- 69 -
Прикрепление микрофиламентов к плазмолемме осуществляется благодаря их связи с её интегральными ("якорными") белками (интегринами) - непосредственно или через ряд промежуточных белков - талин, винкулин и α- актинин (см. рис. 10-9). Помимо этого, актиновые микрофиламенты прикрепляются к трансмембранным белкам в особых участках плазмолеммы. называемых адгезионными соединениями, или фокальными контактами, которые связывают клетки друг с другом или клетки с компонентами межклеточного вещества.
Актин - основной белок микрофиламентов - встречается в моно-мерной форме (G-, или глобулярный актин), которая способна в присутствии цАМФ и Са2+ полимеризоваться в длинные цепи (F-, или фибриллярный актин). Обычно молекула актина имеет вид двух спирально скрученных нитей (см. рис. 10-9 и
13-5).
В микрофиламентах актин взаимодействует с рядом актин-связывающих белков (до нескольких десятков видов), выполняющих различные функции. Некоторые из них регулируют степень полимеризации актина, другие (например, филамин в кортикальной сети или фимбрин и виллин в микроворсинке) способствуют связыванию отдельных микрофиламентов в системы. В немышечных клетках на актин приходится примерно 5-10% содержания белка, лишь около половины его организовано в филаменты. Микрофиламенты более устойчивы к физическим и химическим воздействиям, чем микротрубочки.
Функции микрофиламентов:
(1)обеспечение сократимости мышечных клеток (при взаимодействии с миозином);
(2)обеспечение функций, связанных с кортикальным слоем цито-плазмы и плазмолеммой (экзо- и эндоцитоз, образование псевдоподий и миграция клетки);
(3)перемещение внутри цитоплазмы органелл, транспортных пузырьков и других структур благодаря взаимодействию с некоторыми белками (минимиозином), связанными с поверхностью этих структур;
(4)обеспечение определенной жесткости клетки за счет наличия кортикальной сети, которая препятствует действию деформаций, но сама, перестраиваясь, способствует изменениям клеточной формы;
(5)формирование сократимой перетяжки при цитотомии, завершающей клеточное деление;
(6)образование основы ("каркаса") некоторых органелл (микро-ворсинок,
стереоцилий).
(7)участие в организации структуры межклеточных соединений
(опоясывающих десмосом).
-70 -