Материал: Современные представления о механизмах памяти
Современные представления о механизмах памяти
Введение
В моей курсовой работе речь пойдет о механизмах памяти, выдвинутых гипотезах и экспериментах, также попытаюсь разобраться в роли циркуляции импульсов по кольцевым связям в процессах памяти и условиях запоминания. Но для этого необходимо для начала дать определение понятию «память», а какие виды памяти существуют, рассмотрю в первой главе моей работы уже более подробно.
Памятью называется способность человека сохранять и воспроизводить информацию о ранее произошедших событиях внешнего мира. Различают кратковременную, промежуточную и долговременную память. Кратковременная память характеризуется временем хранения информации от долей секунды до нескольких минут, при этом определяется значимость поступающей информации: если информация важна, то она переходит в другую категорию памяти, если нет, то исчезает. Промежуточная память длится в течение десятков минут или часов и определяет формирование мыслей на определенный период времени. В долговременной памяти информация может сохраняться в течение всей жизни человека, что и обеспечивает запечатление наиболее важных событий.
В основе механизма памяти лежит циркуляция нервных импульсов по кольцевым связям нейронов коры больших полушарий. При формировании долговременной памяти происходит также фиксация, хранение и воспроизведение временных связей в виде белковых молекул.
В запечатлении и извлечении следов памяти участвуют тысячи нейронов коры головного мозга, лимбической системы, таламуса и других нервных центров. Эти следы, как известно, распределены диффузно, но в хранении и извлечении памяти особую роль играют два отдела коры - гиппокамп и височная доля неокортекса.
Для образования долговременной памяти требуется несколько часов. Процесс запечатления следов памяти легко может быть нарушен - например электорошоком, охлаждением мозга или наркозом. Особенно четко эти нарушения выявляются, если воздействовать на гиппокамп. Больные с двусторонними поражениями гиппокампа не способны к длительному запоминанию, хотя опыт, запечатленный до поражения гиппокампа, не страдает.
Деятельность нейронов височной доли тесно связана с хранением и извлечением следов долговременной памяти. При электрическом раздражении височных долей возникают живые картины из прошлого, например, занятия в школе или просмотренный кинофильм. Эти картины очень ярки. Так, если у женщины возникают воспоминания о родах, то она как бы снова переживает все обстоятельства этого события - разговор врача с акушеркой, обстановку в родильной комнате, возбуждение и боль.
Несмотря на то, что височной доле приписывают
функцию извлечения следов памяти, ее не считают хранителем этих следов.
Большинство нейрофизиологов полагают, что память обусловлена деятельность
большого количества нейронов, локализованных диффузно в коре головного мозга, а
также в таких подкорковых образованиях, как лимбическая система, таламус и
гипоталамус.
1. Виды памяти
Виды памяти классифицируют по форме проявления (образная, эмоциональная, логическая, или словесно-логическая), по временной характеристике, или продолжительности (мгновенная, краткосрочная, долгосрочная).
Образная память проявляется формированием, хранением и воспроизведением ранее воспринятого образа реального сигнала, его нервной модели. Под эмоциональной памятью понимают воспроизведение некоторого пережитого ранее эмоционального состояния при повторном предъявлении сигнала, вызвавшем первичное возникновение такого эмоционального состояния. Эмоциональная память характеризуется высокой скоростью и прочностью. В этом, очевидно, главная причина более легкого и устойчивого запоминания человеком эмоционально окрашенных сигналов, раздражителей. Напротив, серая, скучная информация запоминается намного труднее и быстро стирается в памяти. Логическая (словесно-логическая, семантическая) память - память на словесные сигналы, обозначающие как внешние объекты и события, так и вызванные ими ощущения и представления.
Мгновенная (иконическая) память заключается в образовании мгновенного отпечатка, следа действующего стимула в рецепторной структуре. Этот отпечаток, или соответствующая физико-химическая энграмма внешнего стимула, отличается высокой информативностью, полнотой признаков (отсюда и название «иконическая память», т.е. четко проработанное в деталях отражение) действующего сигнала, но и высокой скоростью угасания (хранится не более 100 - 150 мс, если не подкрепляется). Нейрофизиологический механизм иконической памяти, очевидно, заключается в процессах рецепции действующего стимула и ближайшего последействия (когда реальный стимул уже не действует), выражаемого в следовых потенциалах, формирующихся на базе рецепторного электрического потенциала. Продолжительность и выраженность этих следовых потенциалов определяются как силой действующего стимула, так и функциональным состоянием воспринимающих мембран рецепторных структур. Стирание следа памяти происходит за 100-150 мс.
Биологическое значение иконической памяти
заключается в обеспечении анализаторных структур мозга возможностью выделения
отдельных признаков и свойств сенсорного сигнала, распознавания образа.
Иконическая память хранит в себе не только информацию, необходимую для четкого
представления о сенсорных сигналах, поступающих в течение долей секунды, но и
содержит несравненно больший объем информации, чем может быть использован и
реально используется на последующих этапах восприятия, фиксации и
воспроизведения сигналов.
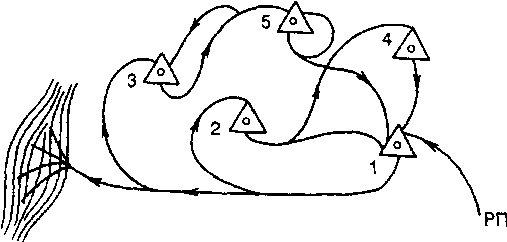
Рис. 1 - Циркуляция возбуждения по цепочке
нейронов (1, 4) и по одному (3, 5) нейрону; РП - рецептивное поле
При достаточной силе действующего стимула иконическая память переходит в категорию краткосрочной (кратковременная) памяти.
Краткосрочная память - оперативная память, обеспечивающая выполнение текущих поведенческих и мыслительных операций. В основе ее лежит повторная многократная циркуляция импульсных разрядов по круговым замкнутым цепям нервных клеток (рис. 1). Кольцевые структуры могут быть образованы и в пределах одного и того же нейрона путем возвратных сигналов, образуемых концевыми разветвлениями аксонного отростка на дендритах этого же нейрона. В результате многократного прохождения импульсов по этим структурам в последних постепенно образуются стойкие изменения, закладывающие основу последующего формирования долгосрочной памяти. В кольцевых структурах могут участвовать не только возбуждающие, но и тормозящие нейроны. Продолжительность краткосрочной памяти составляет секунды, минуты после непосредственного действия соответствующего сообщения, явления, предмета. Реверберационная гипотеза природы краткосрочной памяти допускает наличие замкнутых кругов циркуляции возбуждения как внутри коры большого мозга, так и между корой и подкорковыми образованиями (в частности, таламокортикальные нервные круги), содержащими как сенсорные, так и гностические (обучаемые, распознающие) нервные клетки. Внутрикорковые и таламокортикальные реверберационные круги как структурная основа нейрофизиологического механизма краткосрочной памяти образованы корковыми пирамидными клетками V - VI слоев преимущественно лобных и теменных областей коры большого мозга.
Участие структур гиппокампа и лимбической системы мозга в краткосрочной памяти связано с реализацией этими нервными образованиями функции различения новизны сигналов и считывания поступающей афферентной информации на входе бодрствующего мозга. Реализация феномена краткосрочной памяти не связана с существенными химическими и структурными изменениями в нейронах и синапсах, так как для соответствующих изменений в синтезе матричных (информационных) РНК требуется большее время.
Несмотря на различия гипотез о природе краткосрочной памяти, исходной их предпосылкой является возникновение непродолжительных обратимых изменений физико-химических свойств мембраны, а также динамики медиаторов в синапсах. Ионные токи через мембрану в сочетании с кратковременными метаболическими сдвигами во время активации синапсов могут привести к изменению эффективности синаптической передачи, длящейся несколько секунд.
Превращение краткосрочной памяти в долгосрочную (консолидация памяти) в общем виде обусловлено наступлением стойких изменений синаптической проводимости как результат повторного возбуждения нервных клеток. По данным нейрофизиологии и нейрохимии, в основе долгосрочной памяти лежат процессы синтеза белковых молекул в клетках головного мозга. В консолидации памяти участвует много факторов, облегчающих передачу импульсов по синаптическим структурам (усиленное функционирование определенных синапсов, повышение их проводимости для адекватных импульсных потоков). Одним из таких факторов может служить известный феномен посттетанической потенциации, поддерживаемый реверберирующими потоками импульсов: раздражение афферентных нервных структур приводит к достаточно длительному (десятки минут) повышению проводимости мотонейронов спинного мозга. Это означает, что возникающие при стойком сдвиге мембранного потенциала физико-химические изменения постсинаптических мембран, вероятно, служат основой для образования следов памяти, отражающихся в изменении белкового субстрата нервной клетки.
Определенное значение в механизмах долгосрочной
памяти имеют изменения, наблюдающиеся в медиаторных механизмах, обеспечивающих
процесс химической передачи возбуждения с одной нервной клетки на другую,
например взаимодействие медиаторов с рецепторными белками постсинаптической
мембраны и ионами (Na+, K+, Са2+). Динамика трансмембранных токов этих ионов делает
мембрану более чувствительной к действию медиаторов. Установлено, что процесс
обучения сопровождается повышением активности фермента холинэстеразы,
разрушающей ацетилхолин, а вещества, подавляющие действие холинэстеразы,
вызывают существенные нарушения памяти.
2. Гипотезы о нервных механизмах
памяти
Одной из распространенных химических теорий
памяти является гипотеза Хидена о белковой природе памяти. По мнению автора,
информация, лежащая в основе долговременной памяти, кодируется, записывается в
структуре полинуклеотидной цепи молекулы. Разная структура импульсных
потенциалов, в которых закодирована определенная сенсорная информация в
афферентных нервных проводниках, приводит к разной перестройке молекулы РНК, к
специфическим для каждого сигнала перемещениям нуклеотидов в их цепи. Таким
образом, происходит фиксация каждого сигнала в виде специфического отпечатка в
структуре молекулы РНК. Исходя из гипотезы Хидена, можно предположить, что
глиальные клетки, участвующие в трофическом обеспечении функций нейрона,
включаются в метаболический цикл кодирования поступающих сигналов путем
изменения нуклеотидного состава синтезирующих РНК. Весь набор вероятных
перестановок и комбинаций нуклеотидных элементов обеспечивает возможность
фиксировать в структуре молекулы РНК огромный объем информации: теоретически
рассчитанный ее объем составляет 1015-1020 бит, что значительно перекрывает
реальный объем человеческой памяти. Процесс фиксации информации в нервной
клетке находит отражение в синтезе белка, в молекулу которого вводится
соответствующий следовой отпечаток изменений в молекуле РНК. При этом молекула
белка становится чувствительной к специфическому узору импульсного потока, тем
самым она как бы узнает тот афферентный сигнал, который закодирован в этом
импульсном паттерне. В результате происходит освобождение медиатора в
соответствующем синапсе, приводящее к передаче информации с одной нервной
клетки на другую в системе нейронов, ответственных за фиксацию, хранение и
воспроизведение информации.
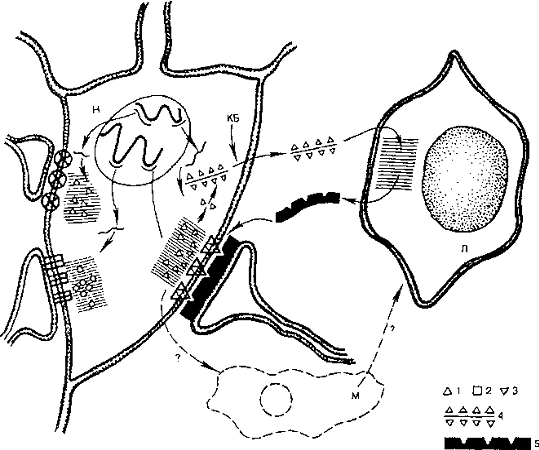
Рис. 2. Иммунохимический механизм формирования
памяти (по И.П. Ашмарину): КБ - катионные белки, открывающие путь комплексам
фрагментов синаптического антигена с РНК через оболочку клетки; Л - клетка глии
(аналог лимфоцитов); М - клетка глии (аналог макрофагов); Н - нейрон; 1, 2, 3 -
участки синаптического антигена (зона синтеза заштрихована); 4 - комплекс
фрагмента синаптического антигена с РНК; 5 - антитела для синапсов. Тонкими
линиями изображены мРНК, толстыми - ДНК хромосом
Возможным субстратом долгосрочной памяти являются некоторые пептиды гормональной природы, простые белковые вещества, специфический белок S-100. К таким пептидам, стимулирующим, например, условнорефлекторный механизм обучения, относятся некоторые гормоны (АКТГ, соматотропин, вазопрессин и др.). Интересная гипотеза об иммунохимическом механизме формирования памяти, предложенная И.П. Ашмариным, основана на признании важной роли активной иммунной реакции в консолидации, формировании долгосрочной памяти. Суть этого представления состоит в следующем: в результате метаболических процессов на синаптических мембранах при реверберации возбуждения на стадии формирования краткосрочной памяти образуются вещества, играющие роль антигена для антител, вырабатываемых в глиальных клетках. Связывание антитела с антигеном происходит при участии стимуляторов образования медиаторов или ингибитора ферментов, разрушающих, расщепляющих эти стимулирующие вещества (рис. 2).
Значительное место в обеспечении нейрофизиологических механизмов долгосрочной памяти отводится глиальным клеткам, число которых в центральных нервных образованиях на порядок превышает число нервных клеток. Предполагают, что на стадии образования и упрочения условного рефлекса в прилегающих к нервной клетке глиальных клетках усиливается синтез миелина, который окутывает концевые тонкие разветвления аксонного отростка и тем самым облегчает проведение по ним нервных импульсов, в результате чего повышается эффективность синаптической передачи возбуждения. В свою очередь стимуляция образования миелина происходит в результате деполяризации мембраны олигодендроцита (глиальной клетки) под влиянием поступающего нервного импульса. Таким образом, в основе долгосрочной памяти могут лежать сопряженные изменения в нервно-глиальном комплексе центральных нервных образований.
Возможность избирательного выключения
краткосрочной памяти без нарушения долгосрочной и избирательного воздействия на
долгосрочную память в отсутствие каких-либо нарушений краткосрочной памяти
обычно рассматривают как свидетельство разной природы нейрофизиологических
механизмов, лежащих в их основе. Косвенным доказательством наличия определенных
различий в механизмах краткосрочной и долгосрочной памяти являются особенности
расстройств памяти при повреждении структур мозга. Так, при некоторых очаговых
поражениях мозга (поражение височных зон коры, структур гиппокампа) при его
сотрясении наступают расстройства памяти, выражающиеся в потере способности
запоминать текущие события или события недавнего прошлого (произошедшие незадолго
до воздействия, вызвавшего данную патологию) при сохранении памяти на прежние,
давно случившиеся события. Однако ряд других воздействий оказывает однотипное
влияние и на краткосрочную и на долгосрочную память. По-видимому, несмотря на
некоторые заметные различия физиологических и биохимических механизмов,
ответственных за формирование и проявление краткосрочной и долгосрочной памяти,
в их природе намного больше общего, чем различного. Их можно рассматривать как
последовательные этапы единого механизма фиксации и упрочения следовых
процессов, протекающих в нервных структурах под влиянием повторяющихся или
постоянно действующих сигналов.
3. Гипотезы о молекулярных
механизмах памяти
Деление памяти на стадии в значительной мере условно, так, как до сих пор неизвестно, действительно ли они объективно независимы или представляют собой искусственно выделенные этапы непрерывного, единого процесса. Наиболее распространено деление памяти на краткосрочную и долгосрочную. Процесс перехода первой во вторую называется консолидацией. Однако недостаточность такого деления очевидна. В самом деле, говоря о краткосрочной памяти, утверждают, что ее продолжительность составляет от долей секунды до десятков минут. Однако даже индивидуальный опыт каждого человека убеждает в том, что это несравнимые интервалы. Попытка воспроизвести изображение или текст через секунду или через десять минут приводит к совершенно различным результатам.
Такая неопределенность ведет к попыткам выделить промежуточные формы памяти. Основанием для деления процесса памяти на временные стадии служит не только степень сохранности информации через определенные промежутки времени, но и данные биохимического анализа. Итак, выделим три стадии памяти:
) минимальная кратковременная память (КПМ);
) относительно ограниченная во времени память (ООП);
) пожизненная долговременная память (ДПП).
Стадия минимальной кратковременной памяти (КПМ длится не более секунды (а, скорее всего, доли секунды). Объем этой памяти достаточен для того, чтобы обеспечить синтез сиюминутного поступления информации от различных органов чувств, произвести быструю обработку этой информации и оценить ее важность для дальнейшего поведения.
Вероятно, эта стадия памяти связана с изменением функционирования синапса, например, с изменением активности Na-, K-, АТФазы. Доказательством этого утверждения является то, что она нарушается различными ингибиторами функций синапса и не изменяется под влиянием ингибиторов синтеза белка и нуклеиновых кислот. Кроме того, очевидно, что время, в течение которого функционирует КПМ, слишком мало для того, чтобы могли произойти сложные биохимические процессы, включающие синтез новых макромолекул.
Память, относительно ограниченная во времени (ООП) энграмма мозг память нейрон. Продолжительность этой стадии памяти составляет от нескольких секунд до нескольких часов, иногда до нескольких суток. Предполагается, что в течение этого времени происходят процессы консолидации. Та информация, которая не подверглась консолидации, стирается и исчезает из памяти.
Механизмы этой стадии памяти разнообразны: активация протеиназ, модифицирующих белки мембран, фосфорилирование белков и некоторых липидов мембран, образование циклических нуклеотидов, включение синтеза нейроспецифических белков и др. Вероятно, все они способны взаимодействовать друг с другом, и набор этих механизмов зависит от характера запоминаемой информации и определяет в конкретных случаях продолжительность ООП. Подтверждается это тем, что в различных экспериментах ООП нарушается разными физическими или химическими агентами.