Материал: Нуклеиновые кислоты
4.2 Вторичная структура ДНК
Структура двойной спирали ДНК была предложена Френсисом Криком и Джеймсом Уотсоном в 1953 году на основании рентгеноструктурных данных, полученных Морисом Уилкинсом и Розалинд Франклин, и "правил Чаргаффа”, согласно которым в каждой молекуле ДНК соблюдаются строгие соотношения, связывающие между собой количество азотистых оснований разных типов. К 1953 году было известно, что ДНК состоит из 4 нуклеотидов, а каждый из них - из одного азотистого основания, 5-углеродного сахара дезоксирибозы и остатка фосфорной кислоты. Не было понятно, как эти части соединяются в молекулы ДНК.
Позже предложенная Уотсоном и Криком модель строения ДНК была доказана, а их работа отмечена Нобелевской премией по физиологии и медицине 1962 г.
Таким образом, двойная спираль правозакрученная,
полинуклеотидные цепи в ней антипараллельны, т.е. если одна из них ориентирована
в направлении 3'→5', то вторая - в направлении 5'→3'. Поэтому на
каждом из концов молекулы ДНК расположены 5'-конец одной цепи и 3'-конец другой
цепи.
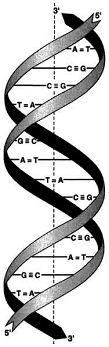
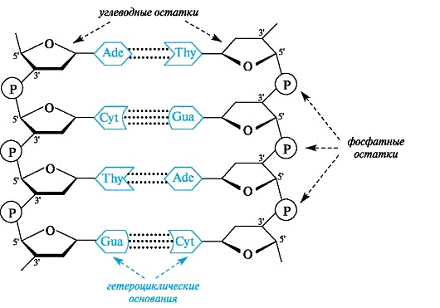
Все основания цепей ДНК расположены внутри двойной спирали, а пентозофосфатный остов - снаружи. Полинуклеотидные цепи удерживаются относительно друг друга за счёт водородных связей между комплементарными пуриновыми и пиримидиновыми азотистыми основаниями А и Т (две связи) и между G и С (три связи). При таком сочетании каждая пара содержит по три кольца, поэтому общий размер этих пар оснований одинаков по всей длине молекулы. Водородные связи при других сочетаниях оснований в паре возможны, но они значительно слабее. Последовательность нуклеотидов одной цепи полностью комплементарна последовательности нуклеотидов второй цепи. Поэтому, согласно правилу Чаргаффа (Эрвин Чаргафф в 1951 г. установил закономерности в соотношении пуриновых и пиримидиновых оснований в молекуле ДНК), число пуриновых оснований (А + G) равно числу пиримидиновых оснований (Т + С).
Комплементарые основания уложены в стопку в сердцевине спирали. Между основаниями двухцепочечной молекулы в стопке возникают гидрофобные взаимодействия, стабилизирующие двойную спираль.
Такая структура исключает контакт азотистых остатков с водой, но стопка оснований не может быть абсолютно вертикальной. Пары оснований слегка смещены относительно друг друга. В образованной структуре различают две бороздки - большую, шириной 2,2 нм, и малую, шириной 1,2 нм. Азотистые основания в области большой и малой бороздок взаимодействуют со специфическими белками, участвующими в организации структуры хроматина.
Т.е. под вторичной структурой понимают пространственную организацию полинуклеотидной цепи. Согласно модели Уотсона-Крика молекула ДНК состоит из двух полинуклеотидных цепей, правозакрученных вокруг общей оси с образованием двойной спирали. Пуриновые и пиримидиновые основания направлены внутрь спирали. Между пуриновым основанием одной цепи и пиримидиновым основанием другой цепи возникают водородные связи. Эти основания составляют комплементарные пары.
Водородные связи образуются между аминогруппой одного
основания и карбонильной группой другого - NH. O=C-, а также между амидным и
иминным атомами азота - NH. N - Например, как показано ниже, между аденином и
тимином образуются две водородные связи, и эти основания составляют
комплементарную пару, т.е. аденину в одной цепи будет соответствовать тимин в
другой цепи. Другую пару комплементарных оснований составляют гуанин и цитозин,
между которыми возникают три водородные связи.
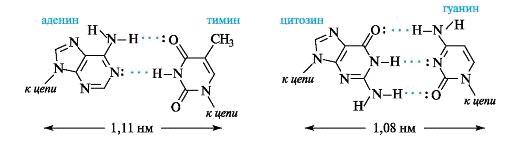
Водородные связи между комплементарными основаниями - один из
видов взаимодействий, стабилизирующих двойную спираль. Две цепи ДНК, образующие
двойную спираль, не идентичны, но комплементарны между собой. Это означает, что
первичная структура, т.е. нуклеотидная последовательность, одной цепи
предопределяет первичную структуру второй цепи.
5. Функции нуклеиновых кислот
5.1 Функции ДНК
. Важнейшая биологическая функция ДНК - генетическая, т.е. ДНК является носителем генетической информации.
. ДНК способна передавать генетическую информацию в ряду поколений посредством и-РНК.
. Регулирует процессы биосинтеза белка,
В клетках эукариот ДНК располагается главным образом в ядре в виде набора хромосом. Бактериальная (прокариоты) ДНК обычно представлена одной кольцевой молекулой ДНК, расположенной в неправильной формы образовании в цитоплазме, называемым нуклеотидом. Генетическая информация генома состоит из генов. Ген - единица передачи наследственной информации и участок ДНК, который влияет на определённую характеристику организма. Ген содержит открытую рамку считывания, которая транскрибируется, а также регуляторные последовательности, например, промотор и энхансер, которые контролируют экспрессию открытых рамок считывания.
Функции хранения и передачи информации обеспечиваются биосинтезом белка, посредством и-РНК и т-РНК.
5.2 Функции РНК
Существуют три типа РНК, каждый из которых выполняет свою особую роль в синтезе белка.
. Матричная РНК переносит генетический код из ядра в цитоплазму, определяя таким образом синтез разнообразных белков.
. Транспортная РНК переносит активированные аминокислоты к рибосомам для синтеза полипептидных молекул.
. Рибосомная РНК в комплексе примерно с 75
разными белками формирует рибосомы - клеточные органеллы, на которых происходит
сборка полипептидных молекул.

Матричная РНК (информационная) представляет собой
длинную одноцепочечную молекулу, присутствующую в цитоплазме. Эта молекула РНК
содержит от нескольких сотен до нескольких тысяч нуклеотидов РНК, образующих
кодоны, строго комплементарные триплетам ДНК. Она переносит генетическую
информацию через мембрану ядра к месту синтеза белка на рибосоме. Переписывание
(перекодирование) информации с ДНК на РНК происходит посредством транскрипции.
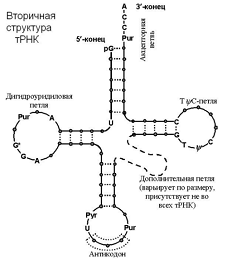
Еще один тип РНК, играющий важнейшую роль в синтезе белка, называют транспортной РНК, поскольку он транспортирует аминокислоты к строящейся молекуле белка. Каждая транспортная РНК специфически связывается только с одной из 20 аминокислот, составляющих белковые молекулы. Транспортные РНК действуют как переносчики специфических аминокислот, доставляя их к рибосомам, на которых происходит сборка полипептидных молекул. Каждая специфическая транспортная РНК распознает "свой" кодон матричной РНК, прикрепившейся к рибосоме, и доставляет соответствующую аминокислоту на соответствующую позицию в синтезируемой полипептидной цепи. Цепь транспортной РНК гораздо короче матричной РНК, содержит всего около 80 нуклеотидов и упакована в форме клеверного листа. На одном конце транспортной РНК всегда находится аденозинмонофосфат (АМФ), к которому через гидроксильную группу рибозы прикрепляется транспортируемая аминокислота. Транспортные РНК служат для прикрепления специфических аминокислот к строящейся полипептидной молекуле, поэтому необходимо, чтобы каждая транспортная РНК обладала специфичностью и в отношении соответствующих кодонов матричной РНК. Код, посредством которого транспортная РНК распознает соответствующий кодон на матричной РНК, также является триплетом и его называют антикодоном. Антикодон располагается примерно посередине молекулы транспортной РНК. Во время синтеза белка азотистые основания антикодона транспортной РНК прикрепляются с помощью водородных связей к азотистым основаниям кодона матричной РНК. Таким образом, на матричной РНК выстраиваются в определенном порядке одна за другой различные аминокислоты, формируя соответствующую аминокислотную последовательность синтезируемого белка.
Рибосомные РНК содержат 3000-5000 нуклеотидов; молекулярная масса - 1 000 000-1 500 000. На долю рРНК приходится 80-85% от общего содержания РНК в клетке. В комплексе с рибосомными белками рРНК образует рибосомы - органоиды, осуществляющие синтез белка. В эукариотических клетках синтез рРНК происходит в ядрышках.
Функции рРНК:
) необходимый структурный компонент рибосом и, таким образом, обеспечение функционирования рибосом;
) обеспечение взаимодействия рибосомы и тРНК;
) первоначальное связывание рибосомы и кодона-инициатора иРНК и определение рамки считывания,
) формирование активного центра рибосомы.
Список использованной литературы
1. Ленинджер А. Основы биохимии: Учебник. - М.: Мир, 1985.
. Тюкавкина Н.А., Бауков Ю.И. Биоорганическая химия: Учебник. - М.: Медицина, 1991.
. Остерман Л.А. Методы исследования белков и нуклеиновых кислот. - М.: Наука, 1981.
. Шабарова 3.А., Богданов А.А., Химия нуклеиновых кислот и их компонентов, М., 1978.
. Зенгер В. Принципы структурной организации нуклеиновых кислот. М., 1987.
6. Сайт: http://ru. wikipedia.org/wiki/%C2%F2%EE%F0%E8%F7%ED%E0%FF_%F1%F2%F0%F3%EA%F2%F3%F0%E0
. Сайт: http://slovari. yandex.ru/нуклеиновые%20кислоты/БСЭ/Нуклеиновые%20кислоты/