Материал: микра от 104 группы
12. Систематика бактерий. Отделы Tenericutes и Mendosicutes. Характеристика и их роль в сельском хозяйстве.
В определителе (Берджи) «Руководство по систематике бактерий Берги» (1984) все прокариотные микроорганизмы были объединены в царство Procaryotae, которое подразделено на четыре отдела — Gracilicutes, Firmicutes, Tenericutes и Мendosicutes. В свою, очередь, отделы делят на классы, порядки, семейства, роды и виды. Микроорганизмы разделены на четыре отдела главным образом на основании наличия или отсутствия клеточных стенок и их вида, а на классы, порядки, семейства, роды, виды — по совокупности морфологических и физиолого-биохимических признаков за пределы вида.
Отдел 3 — Tenericutes.
Отдел объединяет бактерий, не имеющих ригидной клеточной стенки, не синтезирующих пептидогликан. Это плеоморфные организмы, размножающиеся почкованием, фрагментацией и бинарным делением. Микоплазмы могут быть сапротрофами, паразитами и возбудителями болезней животных и растений. Форма клеток может быть сферической или овальной, палочковидной, дисковидной, встречаются и тонкие нити с тенденцией к образованию разветвленных мицелиевидных структур. Считают, что микоплазмы — самые мелкие из всех известных прокариот, имеющих клеточную структуру (0,1—0,25 мкм). Они, подобно вирусам, проходят через бактериологические фильтры, задерживающие обычные бактерии. Размножение микоплазм происходит неправильным делением, что приводит к образованию клеток разной формы и размеров, а также в результате развития в нитях маленьких кокковидных структур — элементарных телец — с их последующим освобождением после разрушения нитей и, наконец, почкованием. Микоплазмы неподвижны. Факультативные анаэробы, хемоорганогетеротрофы. Распространены на растениях и животных, в водоемах, сточных водах и в почве. Микоплазмы объединяют в один класс — Mollicutes (от лат. mollis— мягкий, cutes — кожа). Класс включает порядок Mycoplasmatales, который состоит из трех семейств —Mycoplasmataceae, Acholeplasmataсеае и Spiroplasmataceae. Семейство Mycoplasmataceae представлено родами Mycoplasma и Ureaplasma, виды которых широко распространены в природе (почвах, сточных водах и т. д.). Многие из них сапротрофы и паразиты, в том числе возбудители различных заболеваний человека, животных и растений.
Отдел 4 — Mendosicutes
К отделу Mendosicutes были отнесены прокариоты, обладающие необычной клеточной стенкой, которая не содержит пептидогликана. Клетки имеют форму кокков, палочек и спиралей, а также пирамид, шестилучевой звезды, квадрата, мицелиальных ансамблей и т. д. Они различно окрашиваются по Граму. Эндоспор не образуют; многие виды подвижны. Известны как строгие анаэробы, так и аэробы. Многие встречаются в экстремальных местообитаниях. Представлены классом Archaebacteria. К нему относят прокариот, обладающих уникальными физиологическими, биохимическими свойствами и экологией, резко отличными от остальных прокариот. Так, они отличаются от других бактерий составом и первичной структурой рибосомальных 16S и 5S рРНК, а также транспортных РНК; составом мембранных липидов и образованием однослойной липидной мембраны; составом клеточных стенок (состоят не из пептидогликана, а из других биополимеров — кислых полисахаридов, белков и псевдомуреина); отсутствием сложных жизненных циклов, патогенных и паразитических видов, экзоферментов; способностью использовать только низкомолекулярные органические соединения; жизнеспособностью некоторых видов даже при температуре выше 100 °С и другими признаками. Среди архебактерий выделяют пять основных групп: метанообразующие, аэробные сероокисляющие, анаэробные серовосстанавливающие, галобактерии и термоацидофильные «микоплазмы».
Группа 1 — метаногены. Представлена целым рядом родов, в том числе Methanobacterium, Methanococcus, Methanosarcina и др. Для данной группы характерны палочковидные или кокковидные клетки, подвижные и неподвижные. Спор не образуют. Строгие анаэробы. Облигатные и факультативные хемолитотрофы и хемоорганотрофы. Мезофилы, термофилы, имеются галофильные виды. Необходимо еще раз отметить, что на сегодняшний день архебактерии наряду с эубактериями и эукариотами получили наиболее высокий тасономический статус — домена (Domain). Новые названия доменов, представленных прокариотами, — археи (Archaea) и бактерии (Bacteria) вместо архебактерий и эубактерий, соответственно.
Энергию получают при окислении Н2 с восстановлением СО2 до СН4 либо при использовании уксусной кислоты или метилового спирта с образованием метана и СО2. Метаногены широко распространены в почвах, илах, желудочно-кишечном тракте животных. Нашли они и практическое применение — выделяющийся из отстойников со сточными водами метан собирают и используют как топливо.
Группа 2 — аэробные сероокисляющие архебактерии — представлена родом Sulfolobus. Эти организмы окисляют элементарную серу, используя ее в качестве источника энергии. Факультативные хемолитоавтотрофы. Аэробы. Термофилы, развиваются при температуре 70—75°С, ацидофилы (оптимум рН 3).
Группа 3 — анаэробные серовосстанавливающие архебактерий. В ней выделяют роды Thermoproteus, Thermofilum, Desulfurococcus и др. Представители группы восстанавливают элементарную серу до H2S. Строгие анаэробы. Облигатные и факультативные хемолитотрофы и хемоорганотрофы. Экстремальные термофилы — оптимальная для их развития температура от 85 до 105 °С. Распространены в гидротермальных источниках.
Группа 4 — галобактерии. К ней отнесены роды Halococcus, Halobacterium, Haloarcula. Морфология клеток галобактерии весьма разнообразна — они могут быть палочковидной, кокковидной, квадратной и коробочковидной формы. Галобактерии способны развиваться на средах с высокими концентрациями NaCl (20—25%). Это так называемые экстремальные галофилы. Среди них имеются аэробы и факультативные анаэробы. Необычная физиологическая особенность галобактерий — способность к особому типу фотосинтеза — фотофосфорилированию с участием пурпурной мембраны, в которой содержится пигмент бактериородопсин, поглощающий световую энергию. Кроме того, для них характерно использование ионов Na+ в биоэнергетических процессах. Участвуют в превращении углерода и азота в засоленных почвах, соленых озерах с высокой температурой воды и низким содержанием кислорода, солеварнях и других субстратах.
Группа 5 — термоацидофильные «микоплазмы» — представлена одним родом — Thermoplasma. Это хемоорганотрофы, развивающиеся при высокой температуре (60 °С) и кислотности (рН 1—2). Аэробы. Обнаружены в Японии в горячих источниках.
13. Анаэробное дыхание микроорганизмов с использованием кислорода нитратов и сульфатов. Микроорганизмы, вызывающие эти процессы и продукты восстановления.
При анаэробном дыхании конечным акцептором электронов в электронтранспортной цепи являются неорганические или органические соединения. Например, если конечным акцептором электронов является сульфат, то процесс называют сульфатным дыханием, а бактерии – сульфатвосстанавливающими или сульфатредуцирующими.
Сульфатное
дыхание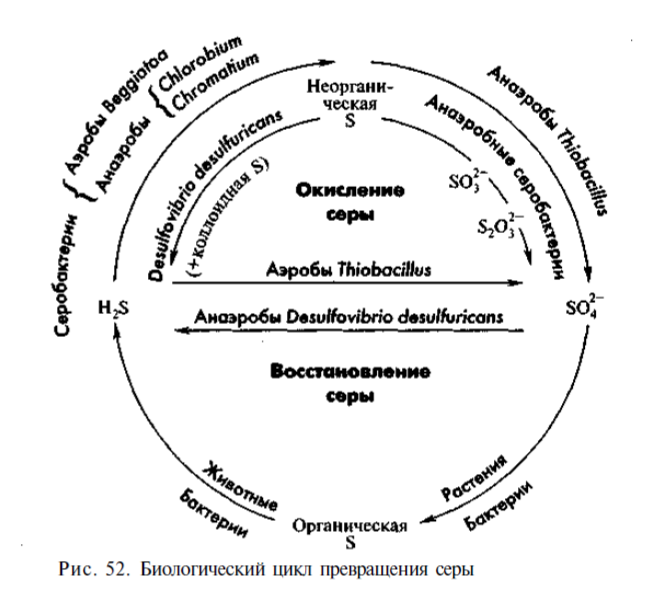
Представители родов |
Краткое описание |
Домен Bacteria |
|
Спорообразующие |
|
Desulfotomaculum Desulfosporosinus |
Грамположительные, разные по морфологии микроорганизмы, образующие эндоспоры. Есть представители, способные как к полному, так и к неполному окислению субстратов. Некоторые — автотрофы, азотфиксаторы, термофилы. Имеют цит b (с), не содержат десульфовиридина |
Бессноровые |
|
Desulfobacterium |
Мелкие палочки с полным окислением субстратов, некоторые — автотрофы |
Desulfococcus |
Кокки, полностью окисляющие субстраты |
Desulfonema |
Нити, передвигающиеся скольжением. Полностью окисляют ацетат |
Thermodesulfobacterium |
Термофильные палочки с неполным окислением субстратов |
Desulfosarcina |
Крупные кокки в кубовидных пакетах с полным окислением ацетата |
Desulfovibrio |
Подвижные вибрионы, содержащие цит с, b и десульфови- ридин. Имеют незамкнутый ЦТК |
Домен Archaea |
|
Archaeoglobus fulgidus |
Архей с нерегулярной формой клеток из морского гидротермального источника. Кроме сульфата, хорошо восстанавливает тиосульфат с глюкозой, формиатом, формами- дом, лактатом, пируватом в качестве доноров электронов. 112 может быть донором при росте с тиосульфатом, но не с сульфатом. Элементарную серу не восстанавливает |
Все сульфатвосстанавливающие бактерии, за исключением представителей двух родов (Desulfotomaculum и Desulfosarcina), имеют клеточную стенку грамотрицательного типа.
Сульфат восстанавливается в соответствии со следующей реакцией:
![]()
Сульфатное дыхание является диссимиляционной сульфатредукцией.
Донорами электронов при сульфатном дыхании могут служить молекулярный водород, органические кислоты, спирты и другие органические соединения. При истощении сульфата и наличии органических веществ сульфатредукторы могут переходить на сбраживание молочной и пировиноградной кислот с образованием ацетата и Н2. Сульфатвосстанавливающие микроорганизмы обычно обитают в морских донных осадках и являются важным звеном глобального круговорота серы. Большая часть сероводорода в природе возникает именно благодаря сульфатному дыханию. Образующийся сульфид служит донором электронов при бактериальном фотосинтезе. В то же время накопление сероводорода в водоемах может вызывать замор рыб. Деятельность сульфатредукторов приводит к коррозии металла и бетона.
Нитратное дыхание
Конечными акцепторами электронов при нитратном дыхании являются нитраты (NO3-) или нитриты (NO2-). Результатом нитратного дыхания является восстановление NO3- или NO2- до газообразных продуктов (NО, N2O, N2). Суммарную реакцию нитратного дыхания, где окисляемым субстратом является глюкоза, а конечным акцептором электронов – нитраты, можно записать следующим образом:
С6Н12О6 + 4NО-3 → 6СО2 + 6Н2О + 2N2 + х(кДж)
Полный процесс денитрификации (диссимиляционная нитротредукция) состоит из четырех реакций восстановления, каждая из которых катализируется специфическими мембраносвязанными редуктазами.
Первый этап: восстановление нитрата до нитрита, катализируют молибденсодержащие ферменты нитратредуктазы:
NО-3 + 2е- + 2Н+ → NО-2 + Н2О
Второй этап: восстановление нитрита до оксида азота, катализируют нитритредуктазы:
NО-2 + е- + Н+ → NО + ОН-
Данные реакции могут протекать только в том случае, когда кислород полностью отсутствует или когда его концентрация незначительна.
Третий этап: восстановление оксида азота до закиси азота, катализируют редуктазы оксида азота:
2NО + 2е- + 2Н+ → N2О + Н2О
Четвертый этап: восстановление закиси азота в молекулярный азот, катализируют редуктазы закиси азота:
N2О + 2е- + 2Н+ → N2 + Н2О
+У денитрифицирующих бактерий, которые являются факультативными анаэробами, функционирует полная электронтранспортная цепь в случае аэробного дыхания, и укороченная – при анаэробном дыхании.
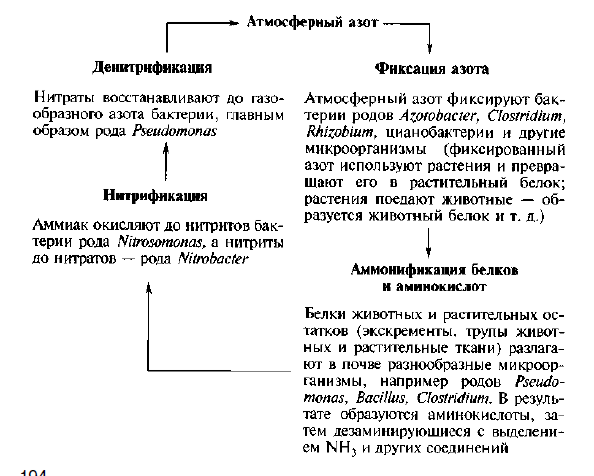
Распространение и роль денитрифицирующих бактерий. Денитрифицирующие бактерии широко распространены в природе. Они принадлежат ко всем основным физиологическим группам: фототрофным, хемолитотрофным, грамположительным и грамотрицательным факультативным анаэробам. Однако в большей степени способность к денитрификации распространена у бактерий родов Bacillus и Pseudomonas.
+Денитрифицирующие бактерии – это обитатели пресных и морских водоемов, почв разного типа, хотя процесс денитрификации у них происходит только в анаэробных условиях. Процесс денитрификации считается вредным для сельского хозяйства, так как доступные для растений нитраты превращаются в недоступный для них молекулярный азот, что приводит к обеднению почвы азотом. Тем не менее, денитрифицирующие бактерии являются важным звеном в круговороте азота в природе, обогащающим атмосферу молекулярным азотом. Кроме того, эти бактерии играют положительную роль в очистке подземных вод и почв от накопившихся в результате деятельности человека (внесение высоких доз удобрений, промышленные стоки) нитратов и нитритов, которые в больших концентрациях токсичны для живых организмов. В связи с этим денитрифицирующие бактерии используют для очистки сточных вод от нитратов.
14. Систематика бактерий. Отдел gracilicutes. Характеристика основных групп грамотр. Бактерий. Значение в природе и сельском хозяйстве.
В последнее время разработаны новые подходы к характеристике микроорганизмов, что оказало огромное влияние на из систематику. В частности определенную ценность имеют методы геносистематики, позволяющие непосредственно охарактеризовать наследственные свойства(генотип) микроорганизмов и Т.О дополнить их описание, которое до последнего времени отражало исключительно структурные и функциональные свойства(фенотип). По соотношениям пар пуриновых и пиримидиновых оснований в молекуле ДНК выделяют генетические различия между группами организмов. Второй метод помогает установить гомологию ДНК при гибридизации пары исследуемых молекул, выделенных из разных микроорганизмов. Если наблюдается высокая степень связывания молекул ДНК(80-90 % И БОЛЕЕ),то можно говорить о гомологии первичной структуры и близком генетическом родстве микроорганизмов(филогенетической связи). Низкая степень гомологии (50%) характеризует достаточно отдаленные связи между микроорганизмами. Особое значение для развития геносистематики имела разработка методов секвенирования, т.е определения последовательности нуклеотидов в нуклеиновых кислотах, в частности в р-РНК. Сравнение таких последовательностей у разных микроорганизмов и построение так называемых фологенетических деревьев, в определенной мере отражающих степень родства между ними, привело к коренным изменениям в систематике.
В систематике иногда используют нумерическую таксономию, предложенную современником Карла Линнея М. Адансоном. В основу такой таксономии положены следующие принципы: равномерность изучаемых признаков организмов; доведение их количества до максимальной величины; выделение каждой таксономической группы по числу совпадающих признаков. После подробного изучения бактерии дают название, которое должно быть выражено 2 словами. Как это требует биномальная номенклатура, предложенная в 18 в Линнеем. Первое слово название рода пишется с прописной буквы(отражает морф. Или физ. Признак, либо фамилию ученого, либо особоый отличительный признак). Второе слово пишется со строчной буквы и обозначает видовое название организма) . названия присваиваются в соответствии с правилами Международного кодекса номенклатуры бактерий. В классификации используют следующие таксономические категории: вид(species), род(genus), семейство( familia), порядок( ordo), класс( classis), отдел(divisio), царство(regnum). В микробиологии часто используют понятие штамм-более узкое, чем вид. Штаммы-культуры того же вида, выделенные из различных природных сред ил из одной и той же среды, но в разное время.
Клон-культура полученная из одной клетки. Совокупность микроорганизмов, состоящую из особей одного вида, называют чистой культурой.
Современные системы классификации микроорганизмов по существу искусственные .
ОТДЕЛ Gracilicutes( от лат. Cutes – кожа, gracilis –тонкий, стройный) К нему относят бактерий(кокки,палочки или нити), имеющих грамотрицательный тип клеточной стенки. Они мб подвижными и неподвижными, эндоспор не образуют. У миксобактерий наблюдается образование плодовых тел и миксоспор. Размножаются бинарным делением, почкованием. В этот отдел входят фототрофные и нефототрофные бактерии, аэробы, анаэробы и факультативные анаэробы. Некоторые виды являются облигатными внутрикл. Паразитами.
ГРУППА 1 СПИРОХЕТЫ
Объединены в порядок Spirochaetales,семейства Spirochaetaceae и Leptospiraceae.
Спирохеты- гибкие, спирально извитые одноклеточные бактерии, представляющие собой очень длинные (5-500 мкм) и тонкие (0,09- 0,75 мкм) клетки с одним или более витками спирали. Клетки состоят из протоплазматического цилиндра. ПЦ и фибриллы покрыты внешней оболочкой. Клетки имеют нуклеотид, мезосомы и др. структуры. Размнлжаются поперечным делением, подвижные. Спор не образуют. Аэробы, факультативные анаэробы или анаэробы. Хемоорганогетеротрофы.
Группа 2-анаробные спиральные и вибриоидные грамотр. бактерии – включает семейство Spirillaceaе. Клетки- жесткие,спирально извитые палочки, подвижные, имеют 1 жгутик или пучок полярно расположенных жгутиков. Аэробы, микроаэрофилы. Хемаорганотрофы. В цитоплазме имеются гранулы поли-В-гидроксимасляной к-ты. Сапротрофы или паразиты. К данному семейству относят роды –Spirillum, Oceanospirillum, Azospirillum, Сampylobacter и Bdellovibrio. Azospirillum-азотфиксирующие.