Материал: микра от 104 группы
7. Процессы получения энергии микроорганизмами. Химизм процессов. Как расходуется полученная энергия микроорганизмами?
Микроорганизмы могут использовать два вида энергии: энергию видимого света и химическую энергию. Фототрофы (водоросли, цианобактерии, пурпурные бактерии и др.) обладают способностью к фотосинтезу. Световая энергия в процессе фотосинтеза улавливается фотоактивными элементами клетки и затем трансформируется в химическую энергию, которая в дальнейшем обеспечивает энергетические потребности микроорганизма.
Химическая энергия, высвобождается в результате окисления кислородом воздуха неорганических соединений (NH3, H2S и др.) и является источником энергии для хемолитотрофов. Энергия, получаемая в процессе окисления органических соединений, является источником энергии для хемоорганотрофов.
Энергия, которая высвобождается в процессе дыхания или брожения, непосредственно не может использоваться микроорганизмами. Для этого свободная энергия преобразуется в другую форму энергии — химическую. Соединением, которое содержит энергию, является АТФ. Также аккумуляторами и переносчиками энергии являются и другие соединения: аденозиндифосфат АДФ, цитозинтрифосфат ЦТФ и др. АТФ часто называют энергетической валютой, из него клетка получает необходимую ей энергию.
Окисление веществ в микробных клетках может происходить различными путями: присоединением к веществу кислорода и отнятием водорода. Окисление происходит также путем переноса электронов от одного вещества к другому. Вещество, теряющее электроны, окисляется (оно называется донором), а присоединяющее электроны - восстанавливается (это акцептор). Перенос водорода осуществляется окислительно-восстановительными ферментами. Акцептором водорода является или кислород воздуха, или любое другое вещество, которое способно восстановиться. В зависимости от того, что является конечным акцептором водорода, микроорганизмы делят на две основные группы:
• аэробные (от греч. аег — воздух) микроорганизмы, окисляющие органические вещества за счет использования кислорода воздуха, который является и акцептором водорода (электронов);
• анаэробные (от греч. ап — частица отрицания + аег — воздух) микроорганизмы, которые в энергетических процессах не используют кислород. Конечным акцептором водорода являются различные соединения, как органические, так и неорганические.
Таким образом, аэробные микроорганизмы окисляют органические вещества до углекислого газа и воды. Если конечным акцептором электронов служит не кислород воздуха, а неорганические соединения (нитраты, сульфаты или карбонаты), то речь идет о анаэробном дыхании (нитратном, сульфатном, карбонатном и др.). К микроорганизмам, способным осуществлять нитратное дыхание, относятся факультативные анаэробы родов Pseudomonas и Bacillus.
В процессе аэробного дыхания последовательно протекают гликолитические реакции, в ходе которых окисляются органические соединения, далее идет комплекс реакций, известных под названием «цикл Кребса», или «цикл трикарбоновых кислот». Затем образуется АТФ и происходит окисление субстрата до воды и углекислого газа. Цикл трикарбоновых кислот имеет большое значение для биосинтеза, так как в соответствии с ним происходит образование соединений, которые легко трансформируются в белки, аминокислоты, липиды, углеводы и другие продукты, необходимые микробной клетке.
Энергетическим материалом для микроорганизмов чаще всего являются углеводы. Сначала проходит гидролиз сложных полисахаридов с образованием моносахаров, чаще всего глюкозы, которая и окисляется. При этом окислении глюкозы освобождается вся энергия молекулы. Этот процесс многоэтапный, он происходит с участием множества разнообразных ферментов, образуется большое количество различных соединений. Но главный промежуточный продукт окисления глюкозы — пировиноградная кислота. Пути расщепления глюкозы до пировиноградной кислоты известны разные, но главным является гликолитический (от греч. glycys — сладкий + lysis — растворение, распад).
Анаэробные микроорганизмы получают энергию в ходе процессов брожения или анаэробного дыхания. Брожение — это дыхание без воздуха, или как установил еще в 1860 г. Л. Пастер, «это жизнь без кислорода». Процессы брожения представляют собой окислительно-восстановительные процессы, в ходе которых происходит образование АТФ в анаэробных условиях. В этом случае акцепторами водорода становятся различные органические соединения, которые в свою очередь являются промежуточными продуктами распада используемого исходного вещества.
Энергетическим материалом при брожении, так же как и при дыхании, чаще служат углеводы, и наиболее используемый из них — глюкоза. Глюкоза трансформируется до пировиноградной кислоты у анаэробов так же, как и у аэробных микроорганизмов, по гликолитическому пути. А вот дальнейшее превращение пировиноградной кислоты иное. У одних микроорганизмов, способных к брожению, она является акцептором водорода и далее восстанавливается в продукт брожения, например молочную кислоту. У других анаэробов пировиноградная кислота преобразуется в различные соединения, которые и являются акцепторами водорода.
Акцептор водорода регенерируется (восстанавливается), а восстановленные органические соединения (которые и являются конечными продуктами брожения) выделяются в субстрат. По основному продукту, накапливающемуся в субстрате, процесс и получает название (спиртовое брожение, молочнокислое брожение и др.). Но, кроме конечных продуктов, в субстрате всегда образуются продукты, окисленные не полностью; это органические вещества, содержащие в себе определенный потенциал энергии. Многие бродильные процессы применяют в промышленности. Но они же могут происходить и в процессах порчи пищевых продуктов.
Некоторые микроорганизмы в анаэробных условиях могут окислять органические вещества с использованием неорганических акцепторов водорода (которые при этом восстанавливаются). Такой способностью обладают, например, денитрифицирующие бактерии, восстанавливающие нитраты до свободного азота, что и называется нитратным дыханием. Сульфатное дыхание происходит при восстановлении сульфатов до сероводорода десульфатирующими бактериями (они используют в качестве акцептора электронов сульфаты).
Получается, что природа акцептора водорода (окислителя) у аэробов и анаэробов разная. Но, с одной стороны, и окисление, и брожение являются процессами, обеспечивающими микроорганизмы энергией, а с другой — промежуточные продукты распада (углеводов) являются материалом для строительства (синтеза белков, нуклеиновых кислот, углеводов, липидов и других соединений, необходимых клетке). Интересно, что с точки зрения выхода энергии дыхание более выгодно для микроорганизмов, чем брожение, в процессе дыхания аэробных микроорганизмов синтезируется больше АТФ, чем у анаэробных в процессе брожения.
8. Анаэробное разложение целлюлозы. Микроорганизмы, принимающие участие в этом процессе. Химизм и его значение в природе.
В анаэробных условиях целлюлозу расщепляют представители родов Bacteroides, Butyrivibrio, Clostridium, Fibrobacter, Thermoanaerobacter и Ruminococcus.
Разложение целлюлозы с помощью микроорганизмов проходит в несколько этапов.
Ферментная система, осуществляющая разложение целлюлозы до глюкозы, носит название целлюлазного комплекса. В его состав входят по меньшей мере три фермента: эндо-β1,4-глюканаза, экзо-β-1,4-глюканаза и β-глюкозидаза.
При анаэробном распаде целлюлозы первоначальный продукт ее гидролиза – глюкоза подвергается сбраживанию, в результате чего образуются различные органические кислоты (уксусная, молочная, муравьиная, масляная, янтарная), этанол, СО2, Н2. Состав продуктов брожения отличается у разных видов микроорганизмов. Примером анаэробного сбраживания целлюлозы являются микробиологические процессы, которые осуществляются в рубце жвачных животных.
Анаэробное сбраживание целлюлозы в рубце жвачных животных
В рубце содержится большое количество микроорганизмов. В состав микробиоты рубца входят строго анаэробные бактерии (10^10 – 10^11 клеток / г ) и анаэробные инфузории, относящиеся к родам Diplodinium и Entodinium (10^4 – 10^6 клеток / г). В разложении полимеров в некоторой степени частично участвуют анаэробные грибы (10^2 – 10^4 зооспор / г). Среди прокариот рубца идентифицировано более 200 видов, принадлежащих к целлюлозолитическим, пектолитическим, молочнокислым, маслянокислым, пропионовокислым и метаногенным бактериям.
Общий состав микробиоты рубца довольно стабилен и разнообразен; ни один вид микроорганизмов не составляет более 3 % общей численности микробных клеток. Микроорганизмы рубца расщепляют целлюлозу и другие сложные углеводы, присутствующие в поглощенном животном корме, образуя жирные кислоты и газы.
9. Применение микроорганизмов-антагонистов и антибиотических веществ для защиты растений от болезней и их профилактика. Использование микробных биопрепаратов для борьбы с насекомыми-вредителями с/х культур.
Известно, что болезни растений распространены широко и причиняют существенный вред сельскому хозяйству. Для борьбы с ними используют химические средства, а также более безопасные для окружающей среды биологические методы.
Освобождению почвы от фитопатогенных организмов способствует усиление размножения в ней микробов-антагонистов по отношению к возбудителям тех или иных заболеваний. Например, после посева люцерны почва очищается от возбудителя вертициллезного вилта хлопчатника (VerUcUlium dahliae). Очевидно, это объясняется не только тем, что корневая система люцерны выделяет в почву алкалоиды, угнетающие многие микроорганизмы, но и тем, что она стимулирует размножение в почве антагонистов возбудителя вертициллеза. Подобными свойствами обладают и растения рапса, промежуточная культура которого может быть использована на юге между посевами других культур.
Возделывание некоторых растений, например клевера и вики, способствует освобождению почвы от возбудителя сибирской язвы Bacillus anthracis: житняк, картофель, наоборот, благоприятствуют размножению этой бактерии. Таким образом, в принципе возможна борьба с болезнетворными микробами почвы путем введения в севооборот тех или иных растений. Однако для широкого практического применения этого приема необходима его экспериментальная доработка.
Внимание микробиологов привлекает использование микробов-антагонистов для лечения растений. На грибах-паразитах нередко паразитируют другие грибы, так называемые грибы-паразиты второго порядка. Так, на мучнисторосяных грибах паразитирует пикнидиальный гриб Cicinnobolus cesati; на возбудителе бурой ржавчины пшеницы Puccinia triticina — также пикнидиальный гриб Dar- luca /Пит. Эксперименты с грибами-паразитами второго порядка, которых наносили в виде водных суспензий на поверхность растений в профилактических целях или при борьбе с заболеваниями, дали обнадеживающие результаты. Однако для профилактических целей перечисленные микроорганизмы пока не применяют.
Хороший эффект дают культуры микробов-антагонистов при обработке семян, зараженных фитопатогенами, или при внесении на поверхность вегетирующих растений, а также в зараженную почву. Микроб-антагонист, уничтожая вредителя, не причиняет вреда растению-хозяину. Микробы-антагонисты угнетают фитопаразитов не только в зоне корня. Вырабатываемые ими антибиотики проникают в ткани растений, повышая их устойчивость к возбудителям болезней.
Я. /7. Худяков (1935) выделил бактерии рода Pseudomonas, лизирующие мицелий фитопатогенных грибов Sc/erotinia и Botry'tis. Этих микробов-антагонистов успешно использовали в полевых опытах для борьбы с фузариозом пшеницы, льна и т. д. Культурой Pseudomonas бактеризовали семена растений. Оздоровлению сеянцев и саженцев сосны способствовало применение Н. Л. Красильниковым миколитических бактерий для борьбы с фузариозом. Как уже было отмечено, культура Azotobacter chroococcum предупреждает заболевания сельскохозяйственных растений, вызываемые рядом грибов, например Alternaria.
Успешно бороться с мучнистой росой крыжовника, вызываемой грибом Sphaerotheca morsuvae, позволяет опрыскивание растений настоем навоза. Это стимулирует размножение микроорганизмов на поверхности растения. В составе эпифитной микрофлоры находятся бактерии-антагонисты, которые после такого опрыскивания начинают размножаться.
Микробов-антагонистов, вероятно, можно использовать не только против возбудителей болезней растений, но и против растений-паразитов культурных растений. Так, положительные результаты были получены при борьбе с заразихой арбуза (Orobanche aegipty- аса)1 с использованием патогенного для заразихи гриба Fusarium orobanches. Чистая культура указанного гриба, размноженная на питательном субстрате (кукурузная мука и г. д.), предложена для практического применения.
Не исключена возможность подбора микробных культур, действующих как гербициды на определенные группы сорных растений.
Культуры некоторых грибов-антагонистов применяют в борьбе с почвенной инфекцией. Установлено, что грибы рода Trichoder- та выделяют токсичные вещества, поражающие микробов-фитопаразитов. При внесении в почву культуры Trichoderma lignorum существенно уменьшается увядание хлопчатника, пораженного Verticillium albo-atrum, снижаются грибные заболевания картофеля и других сельскохозяйственных культур. Вносят культуру указанного гриба в почву при посеве растений. На основе культуры Trichoderma //- gnorum готовят препарат триходермин.
Кратко остановимся на технике использования микробов-антагонистов. Для обеззараживания семена опрыскивают культурой микроорганизмов, разведенной в воде. Стерилизуется не только поверхность семени, но и зона корня, куда переходят микроорганизмы и начинают там размножаться. При высадке рассады и саженцев их корни смачивают взвесью в воде соответствующих микробов-антагонистов. Водную взвесь микробов можно использовать и для опрыскивания надземных частей поврежденных растений, а также для профилактических целей.
Препараты для борьбы с почвенной инфекцией типа триходермина вносят в почву при посеве. Однако пока микробы-антагонисты систематического применения в сельском хозяйстве не получили.
Широко используют микробиологический метод борьбы с фызунами — домашними мышами, полевками, крысами. Известно несколько культур микроорганизмов, вызывающих у грызунов кишечные заболевания, напоминающие брюшной тиф. Для человека и домашних животных данные микроорганизмы безопасны.
Впервые бактерию мышиного тифа — Bacterium typhimurium — выделил в 1892 г. в Германии Ф. Леффлер. Позднее С. С. Мережковский, Б. Л. Исаченко и другие ученые обнаружили ряд близких форм микроорганизмов. Эти бактерии относятся к роду Salmonella (семейство Enterobacteriaceae), они патогенны для человека и животных.
10. Размеры, форма, структурная организация и химический состав бактериальной клетки. Грамположительные и грамотрицательные бактерии. Значение окраски по грамму для диагностики микроорганизмов.
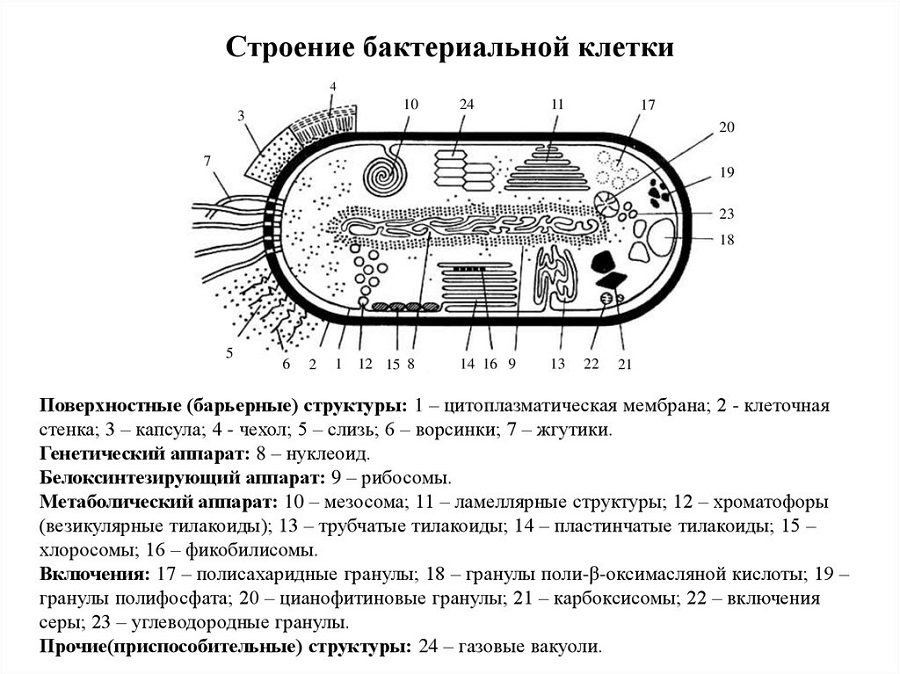
Капсулы. Клетки большинства бактерий окружены слизистым слоем, расположенным поверх клеточной стенки, — капсулой (рис. 15). Встречаются макрокапсулы с толщиной слоя 0,2 мкм, микрокапсулы со слоем менее 0,2 мкм, а также слизистый слой и растворимая слизь. По химическому составу капсулы большинства бактерий можно разделить на два типа. Одни представлены полисахаридами, другие — полипептидами. Однако существуют и капсулы с высоким содержанием липидов, гетерополисахаридов и других веществ. Капсулы содержат до 98% воды, что составляет дополнительный осмотический барьер, они также защищают клетку от механических повреждений и высыхания.
Жгутики. Существуют два основных типа подвижных бактерий: скользящие и плавающие. Скольжение наблюдается у миксобактерий, серных бактерий, цианобактерий и др. Эти организмы способны скользить по твердой поверхности в результате волнообразных сокращений клетки. Плавающие палочковидные бактерии передвигаются при помощи особых нитевидных структур — жгутиков. Так двигается большинство спирил, энтеробактерий, псевдомонад. Кокки, за исключением отдельных видов, жгутиков не имеют. Бактерию с одним жгутиком называют монотрихом; с пучком жгутиков на одном конце клетки — лофотрихом; на обоих концах —амфитрихом; со жгутиками, расположенными по всей поверхности клетки, — перитрихом. В строении жгутика можно выделить нить, крюк и базальное тельце, расположенное под цитоплазматической мембраной.
Движение, ориентированное относительно направления действия какого-либо фактора, носит название таксиса. В зависимости от природы внешнего фактора, под воздействием которого происходит движение, различают хемотаксис, аэротаксис, фототаксис, термотаксис, магнетотаксис и вискозитаксис. Хемотаксис — это движение бактерий в определенном направлении относительно того или иного источника химического соединения — эффектора. Химические соединения для каждой бактерии можно подразделить на вещества, не вызывающие таксисов, и на эффекторы — аттрактанты и репелленты. Аттрактанты — соединения, привлекающие организмы, репелленты — вещества, их отпугивающие. Аттрактанты большей частью представлены пищевыми субстратами, а репелленты — ядовитыми соединениями. Аэротаксис связан с разницей содержания в среде кислорода, а термотаксис — с разницей температур. При фототаксисе условием направленного движения бактерий служит различие в интенсивности освещения. Фототаксис обнаруживается прежде всего у фототрофных бактерий. Магнетотаксис — это способность бактерий двигаться по силовым линиям магнитного поля Земли. В клетках этих организмов имеются включения магнетита, выполняющие функцию магнитной стрелки. Если в Северном полушарии магнетобактерии плывут в направлении Северного полюса, то в Южном — в направлении Южного. Вискозитаксис — это реакция бактерий на изменение вязкости раствора: они способны плыть в направлении ее увеличения или снижения. Считают, что таксисы можно рассматривать как элементарную форму поведения бактерий.
Фимбрии и пили. Кроме жгутиков, клетки бактерий могут иметь длинные, тонкие и прямые нити — фимбрии. Фимбрии значительно короче и тоньше жгутиков, но более многочисленны. Фимбрии первого типа характерны для многих бактерий, в связи с чем их называют фимбриями общего типа. Считают, что фимбрии данного типа служат органами прикрепления
Большой интерес представляют фимбрии второго типа, так называемые половые фимбрии, или F - пили, имеющие внутри канал, через который передается генетический материал от одной клетки к другой при конъюгации бактерий. Половые пили представляют собой белковые цилиндры толщиной 8,5—9,5 нм и длиной до 1,1 мкм. На поверхности клеток некоторых бактерий, близких к псевдомонадам, обнаружены своеобразные структуры, так называемые шипы. Они представляют собой полые цилиндры длиной 1—3 мкм и толщиной около 65 нм. Шипы состоят из белка спинина. Бактерии с шипами обычно неподвижны.