Материал: микра от 104 группы
Окрашивание бактерий по Граму открыта Грамом в 1884 г., позволяющий дифференцировать бактерии. После окраски генцианвиолетом и обработки раствором иода клетки одних видов бактерий обесцвечиваются спиртом, других — остаются окрашенными в сине-фиолетовый цвет. По данному признаку бактерии разделяют на окрашивающиеся по Граму — грамположительные и не окрашивающиеся — грамотрицательные. Большинство Грам (+) бактерий имеют однослойную клеточную мембрану, без внешней мембраны, присущей грамотрицательным бактериям. Исключением является тип Deinococcus-Thermus.
Главным структурным компонентом клеточных стенок большинства исследованных бактерий служит пептидогликан, или муреин. Пептидогликан придает клеточной стенке ригидные свойства, благодаря чему бактериальная клетка способна сохранять форму. У грамположительных бактерий клеточная стенка состоит главным образом из многослойного пептидогликана, с которым соединены вторичные полимеры — тейхоевые кислоты. У грамположительных бактерий пептидогликан обычно составляет 40—60% сухой массы клеточной стенки, у некоторых видов — 80—90%. Отличительная особенность клеточной стенки грамотрицательных бактерий — наличие так называемой наружной мембраны. Наружная мембрана состоит из фосфолипидов, липополисахарида (ЛПС), липопротеина (ЛП) и белков. Толщина клеточной стенки колеблется от 20 до 100 нм и более и составляет около 20% сухого вещества бактериальной клетки. Клеточная стенка относительно проницаема для крупных молекул.

Цитоплазматическая мембрана. К клеточной стенке бактериальной клетки тесно прилегает внешний слой цитоплазмы — цитоплазматическая мембрана. Цитоплазматическая мембрана бактерий служит главным барьером между цитоплазмой клетки и окружающей внешней средой. При разрушении цитоплазматической мембраны бактериальная клетка погибает. В основе цитоплазматической мембраны лежит билипидный слой. Липиды составляют 15—50% сухой массы цитоплазматической мембраны. Основная масса мембранных липидов (70—90%) бактерий представлена фосфолипидами. Около 50% поверхности цитоплазматической мембраны составляют мембранные белки. Они полностью или частично погружены в липидный бислой, некоторые белки располагаются на его поверхности. Общая толщина мембраны составляет приблизительно 7—8 нм. Нередко мембрана дает внутрицитоплазматические впячивания (инвагинации), приводящие к образованию особых структур — мезосом, или нуклеоидосом.
Мезосомы — это мембранные системы, состоящие из трубочек, пузырьков и пластинок. Наиболее обычный тип мезосом — мембранные кольцевые впячивания цитоплазматической мембраны, расположенные в зоне образования клеточной перегородки в клетках бактерий при делении. Предполагается, что они ответственны за специализированные процессы обмена веществ и репликацию ДНК.
Цитоплазма. Под цитоплазматической мембраной у бактерий находится цитоплазма. Это коллоидная система, состоящая из воды, белков, жиров, углеводов, минеральных соединений и других веществ, соотношение которых варьирует в зависимости от вида бактерий и их возраста. Цитоплазма бактерий содержит различные структурные элементы — внутрицитоплазматические мембраны, генетический аппарат, рибосомы и включения; остальная часть ее представлена цитозолем.
Цитозоль — это фракция цитоплазмы, которая имеет гомогенную консистенцию и состоит главным образом из белковых макромолекул (растворимых РНК, ферментных белков, продуктов и субстратов различных реакций). Цитозоль служит поддерживающей средой для клеточных гранул. По структуре цитоплазма мелкогранулярная, состоит из цитоплазматических гранул диаметром 10—20 нм. В цитоплазме находятся рибосомы — частицы, состоящие из РНК (60%) и белка (40%) и имеющие форму круглых или несколько удлиненных структур диаметром около 20 нм. Рибосомы совместно с молекулами РНК и ДНК участвуют в синтезе белка не как изолированные частицы, а в виде агрегатов, называемых полирибосомами, или полисомами. В клетках цианобактерий присутствуют так называемые тилакоиды — внутриклеточные мембранные фотосинтезирующие структуры, содержащие хлорофилл и каротиноиды, при помощи которых осуществляется фотосинтез. Особые светособирающие пигменты — фикобилины, находятся в специальных структурах, фикобилисомах, располагающихся на поверхности тилакоидов. У пурпурных серобактерий фотосинтезирующие пигменты (бактериохлорофилл и каротиноиды) локализованы в хроматофорах, составляющих от 40 до 50% массы клетки. Мембранные структуры хроматофора имеют вид трубочек и пузырьков диаметром 20—100 нм. Трубочки и пузырьки образуют в клетке сложную мембранную сеть и на многих участках сохраняют связь с цитоплазматической мембраной. У зеленых бактерий светособирающие пигменты, участвующие в фотосинтезе, содержатся в особых структурах, называемых хлоросомами.
Включения. В цитоплазме клеток бактерий часто содержатся гранулы различной формы и размеров. Их присутствие нельзя рассматривать как постоянный признак микроорганизма, обычно они в значительной степени связаны с физическими и химическими условиями среды обитания. Примером подобных включений служат газовые вакуоли —аэросомы. Они образованы скоплениями газовых пузырьков, заполненных газом, состав которого соответствует таковому окружающей среды. Как резервные в клетках бактерий могут накапливаться питательные вещества, состоящие из полисахаридов — гранулы гликогена или гранулезы (близкого к амилопектину полисахарида). При недостаточном поступлении углеродсодержащих веществ в питательную среду гранулы гликогена или гранулезы постепенно исчезают из клеток бактерий. Многие бактерии в качестве резервного вещества синтезируют поли-B-гидроксимасляную кислоту (ПОМ) (полиэфир-B-оксимасляной кислоты) данного вещества. У некоторых видов бактерий в клетках накапливаются гранулы жира и волютина. Волютиновые гранулы, называемые еще метахроматическими гранулами, состоят преимущественно из полифосфатов и служат местом запасания фосфора. В клетках серных бактерий встречаются включения серы, которая образуется в результате окисления сероводорода. В клетках цианобактерий, ряда других фототрофных и хемолитоавтотрофных бактерий обнаружены ромбовидные или шестигранные включения, названные карбоксисомами, или полиэдральными телами. Магнитосомы — структуры, представляющие собой частицы Fe3O4, окруженные белковой мембраной, присутствуют в клетках бактерий, обладающих магнитотаксисом, т. е. перемещающихся вдоль линий магнитного поля
Нуклеоид. В цитоплазме бактериальных клеток расположена структура, эквивалентная ядру и называемая нуклеоидом. Нуклеоиды бактерий содержат ДНК, молекулярная масса которой колеблется от 0,45х109 у микоплазм, до 2,9х109 у спорообразующих бактерий и энтеробактерий, что значительно меньше молекулярной массы ДНК эукариот. Установлено, что бактериальная ДНК по форме представляет собой свернутую в кольцо нить длиной 1,1—1,6 мм, называемую также бактериальной хромосомой. Нуклеоид бактерий — основной носитель информации о свойствах клетки и основной фактор передачи этих свойств потомству. Кроме нуклеоида, в цитоплазме бактериальной клетки могут находиться в сотни раз более короткие кольцевые нити ДНК — так называемые внехромосомные факторы наследственности, получившие название плазмиды. Как выяснено, плазмиды не всегда имеются у бактерий. Однако их присутствие обеспечивает дополнительные, полезные для организма свойства, в частности связанные с размножением, устойчивостью к лекарственным препаратам, болезнетворностью и др.
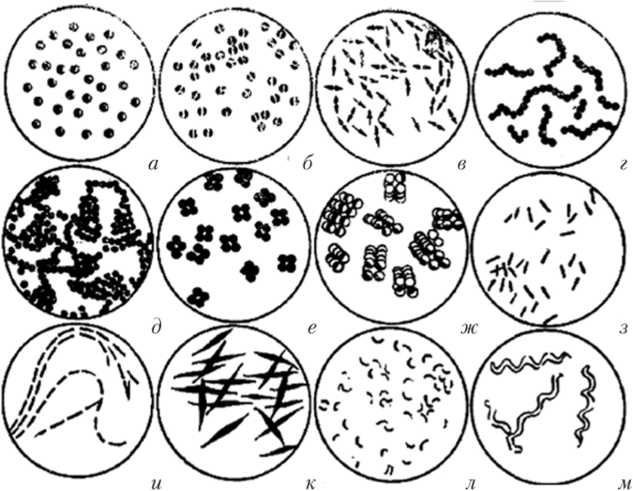
Рис. 2.1. Формы бактерий:
а — кокки; б, в — диплококки; г — стрептококки; д — стафилококки; е — тетракокки; ж — сардины; з — палочковидные бактерии; и — стрептобактерии; к — клостридии; л — вирионы; м — спирохеты
Клетки бактерий очень малы. Их измеряют в микрометрах (мкм), а детали тонкой структуры — в нанометрах (нм). Кокки обычно имеют диаметр около 0,5—1,5 мкм. Ширина палочковидных (цилиндрических) форм бактерий в большинстве случаев колеблется от 0,5 до 1 мкм, длина — от 2 до 10. Мелкие палочки обычно бывают шириной 0,2—0,4 мкм, длиной — 0,7—1,5. Среди бактерий встречаются и настоящие гиганты, длина которых достигает десятков и даже сотен микрометров.
Сферические формы (кокки) - шаровидные бактерии размером 0,5 - 1,0 мкм; по взаимнму расположению клеток различают микрококии, диплококки, стрептококки, тетракокки, сарцины и стафилококки.
Микрококки (.монококки) (от лат. mikros — малый): деление клеток простое, как правило, это сапрофиты, патогенные для человека не обнаружены (рис. 2.2).
Диплококки (от лат. diplos — двойной): деление происходит с образованием пар, клетки имеют бобовидную или ланцетовидную форму (рис. 2.3).
Стрептококки (от лат. streptos — цепочка): деление происходит с образованием цепочек, т.е. размножающиеся клетки сохраняют связь и нередко напоминают бусы. Известны стрептококки, патогенные для человека, которые способны вызывать такие заболевания, как ангина, гнойные воспаления и др. (рис. 2.4).
Стафилококки (от лат. staphyle — гроздь винограда): образующиеся при делении клетки располагаются скоплениями (кистеобразные кластеры), они действительно похожи на гроздья винограда. Стафилококки вызывают более 100 различных заболеваний человека (рис. 2.5).
Тетракокки (от лат. tetra — четыре): клетки делятся с образованием объемных тетрад. В основном это сапрофиты, патогенных для человека почти нет.
Сарцины (от лат. sarcina — связка, тюк): деление клеток происходит с образованием компактно расположенного сочетания клеток из 8, 16 единиц и более. Очень распространены в воздухе, некоторые являются условно-патогенными (рис. 2.6).
Бактерия — это термин (от греч. hakteria — палочка), который давно применяют для всех прокариот. Этим же термином называют неспорообразующие палочковидные бактерии (цилиндрические). Спорообразующие палочки называются бациллы (от лат. bacillus — палочка).
Палочковидные бактерии бывают длинными: более 3—10 мкм (например, бактерии рода Clostridium), короткими: 1,5—3,0 мкм (многие возбудители кишечных заболеваний, в том числе Escherichia coll) и очень короткими: менее 1,0 мкм — в виде коккобактерий (например, возбудитель бруцеллеза Brucella melitensis). Интересно, что концы палочковидных бактерий бывают закругленные, заостренные, обрезанные и даже вдавленные. Палочки могут иметь яйцевидную форму. Условно по толщине палочки делят на тонкие (0,2—0,4 мкм) и толстые (0,7—1,5 мкм).
По взаимному расположению клеток палочковидных бактерий (рис. 2.7) выделяют:
• монобактерии — клетки располагаются одиночно;
• диплобактерии — клетки располагаются попарно;
• стрептобактерии — клетки располагаются цепочкой.
Извитые (спиралевидные) бактерии подразделяются но количеству и характеру завитков и диаметру клеток следующим образом:
• вибрионы (от лат. vibrio — изгибаюсь) — имеют один изгиб, или форму запятой (например, возбудитель холеры) (рис. 2.8);
• спириллы (от лат. spira — завиток) — это клетки, имеющие достаточно большой диаметр и два-три завитка;
• спирохеты (от лат. spira — завиток + греч. chaite — гребень, грива) — клетки с большим числом мелких витков (рис. 2.9).
11. Симбиотическая азотфиксация. Особенности клубеньковых бактерий, их роль в накоплении азота в симбиозе с бобовыми растениями в почве. Биопрепарат «Ризоторфин», основанный на использовании клубеньковых бактерий. Применение и эффективность.
 Симбиотические
азотфиксирующие микроорганизмы выделены
М. Бейеринком в 1888 г. из корневых клубеньков
(бородавчатых наростов) бобовых растений.
Микроорганизмы назвали клубеньковыми
бактериями, и было установлено, что они
вызывают образование клубеньков, в
которых осуществляется фиксация азота
атмосферы. Бактерии в клубеньках питаются
органическими соединениями, синтезированными
растением, а растение получает из
клубеньков связанные соединения азота.
Симбиотические
азотфиксирующие микроорганизмы выделены
М. Бейеринком в 1888 г. из корневых клубеньков
(бородавчатых наростов) бобовых растений.
Микроорганизмы назвали клубеньковыми
бактериями, и было установлено, что они
вызывают образование клубеньков, в
которых осуществляется фиксация азота
атмосферы. Бактерии в клубеньках питаются
органическими соединениями, синтезированными
растением, а растение получает из
клубеньков связанные соединения азота.
Клубеньковые бактерии представляют собой грамотрицательные, от коротких до среднего размера палочки (0,5—0,9 мкм шириной, 1—3 мкм длиной), подвижные, монотрихи с полярным или субполярным расположением жгутиков или перитрихи, аэробы. Спор не образуют. Источником азота для клубеньковых бактерий служат различные соединения — соли аммония и азотной кислоты, многие аминокислоты, пуриновые и пиримидиновые основания и т. д. Клубеньковые бактерии используют разнообразные углеводы, в том числе и некоторые полисахариды (декстрин, гликоген). При усвоении углеводов в процессе жизнедеятельности некоторых видов образуются кислоты. Бактерии потребляют многие органические кислоты и многоатомные спирты. Фосфор клубеньковые бактерии усваивают из минеральных и органических соединений; калий, кальций и другие элементы получают из неорганических веществ. Клубеньковым бактериям нужны также соединения железа, некоторые микроэлементы (молибден и др.)
Бактероиды. Клетки клубеньковых бактерий, перешедшие в цитоплазму растительных клеток, растут, делятся, а затем трансформируются в своеобразные образования — бактероиды. Этим заканчивается процесс инфицирования — приблизительно через три-четыре недели после заражения. Бактероиды в 3—5 раз больше по размерам, чем обычные клетки, причем их форма меняется в зависимости от вида бобового растения от шаровидной и грушевидной до вильчатой и ветвистой. Бактероиды не делятся, они составляют до 50% массы клубенька. Ткань клубенька, заполненная бактероидами, обычно приобретает красноватую окраску благодаря пигменту леггемоглобину, родственному гемоглобину. Обычно такая окраска характерна для клубеньков, активно фиксирующих азот. Бактероиды (лат. Bacteroides) — род грамотрицательных анаэробных бактерий, наиболее типичные нормальные обитатели кишечника человека, составляющие около половины всей его микрофлоры.
Клубеньки, образовавшиеся при инфицировании неактивными клубеньковыми бактериями, содержат мало леггемоглобина и имеют зеленоватый цвет. Клубеньки, образованные активными штаммами, окрашены в розовый цвет. Кроме того, клубеньки неодинаково распределены по корневой системе растений. Активные расы клубеньковых бактерий образуют многочисленные клубеньки на главном корне, а на боковых их бывает мало. По мере старения и дегенерации клубеньки отмирают.
Эффективность азотфиксации симбиотической ассоциации бобовое растение – клубеньковые бактерии определяется наличием у клубеньковых бактерий целого комплекса симбиотических признаков:
вирулентности – способности клубеньковых бактерий входить в контакт с корневой системой бобовых растений, проникать в ткани корня, размножаться в них и индуцировать образование клубеньков;
азотфиксирующей активности – способности связывать молекулярный азот атмосферы при помощи специальной ферментативной системы и превращать его в ионы аммония;
эффективности – способности увеличивать урожай и содержание белка у бобового растения-хозяина за счет передачи растению фиксированного азота и синтезированных биологически активных веществ;
конкурентоспособности – способности внесенного в почву определенного штамма клубеньковых бактерий образовывать клубеньки в присутствии других штаммов того же вида;
специфичности – способности вступать в эффективный симбиоз со строго определенным набором сортов и видов бобовых растений.
Биопрепарат ризоторфин на основе клубеньковых бактерий рода Rhizobium и Bradyrhizobium.
Вскоре после того как М. Бейеринк (1888) изолировал клубеньковые бактерии бобовых растений, возникла идея использовать эти бактерии для улучшения образования клубеньков и усиления фиксации атмосферного азота. Впервые препарат клубеньковых бактерий под названием «нитрагин» был приготовлен в 1896 г. в Германии Ф. Ноббе и Л. Гильтнером.
Это грамотрицательные почвенные бактерии, способные к фиксации азота. Вступают в симбиотические отношения с представителями семейства бобовых, Бактерии рода колонизируют клетки корня растения, образуя корневые клубеньки, и в условиях с пониженным содержанием кислорода способны преобразовывать атмосферный азот в аммиак, обеспечивая растению доступ к органическому азоту в форме глутамина или уреидов. В обмен растение поставляет бактериям сахара, образовавшиеся в ходе фотосинтеза, и обеспечивает им анаэробные условия.
Использование препаратов клубеньковых бактерий для заражения семян бобовых растений совершенно необходимо, когда в данной местности вводят новые культуры бобовых и в составе флоры нет перекрестно заражающихся с ними растений. Такая потребность возникла в нашей стране при возделывании соевых бобов в новых зонах.
В целесообразности применения инокуляции для новых культур бобовых растений, а также на вновь осваиваемых сельскохозяйственных угодьях нет сомнения. Значительно труднее решить вопрос о старопахотных, хорошо окультуренных почвах, на которых уже давно возделывают определенные виды бобовых растений. Для ответа на этот вопрос были поставлены многочисленные опыты. В европейской части России массовые опыты с инокуляцией разных бобовых культур были проведены Е. Н. Мишустиным и В. В. Бернардом. В большинстве случаев инокуляция дала заметное увеличение урожая. Наилучший эффект отмечался на кислых почвах.
Объяснить такое действие заражения бобовых растений культурой Rhizobium или Bradyrhizobium на давно освоенных почвах, имеющих в составе микрофлоры клубеньковые бактерии, можно следующим образом. Во-первых, в природных условиях может происходить перекрестное заражение, т. е. высеваемые бобовые растения заражаются клубеньковыми бактериями близких групп растений. В таких случаях клубеньки хотя и образуются, но функционируют неполноценно. В то же время при искусственной инокуляции в корень бобового растения проникает активная раса Rhizobium или Bradyrhizobium, нанесенная на высеваемые семена. Во-вторых, клубеньковые бактерии, имеющиеся в почве, не занятой бобовыми растениями, существуют как обычные сапротрофы. Нередко вследствие ряда причин почва оказывается неблагоприятной средой для клубеньковых бактерий. Их количество существенно уменьшается, а активность снижается. Кислые почвы, например, отрицательно влияют на азотфиксирующую способность клубеньковых бактерий, и при сапротрофном существовании происходит существенное снижение их ценных свойств.
Бактеризация не только увеличивает урожай бобовых растений, но и улучшает его качество. В растениях, зараженных активными расами клубеньковых бактерий, значительно повышается количество белка и витаминов группы В. Поскольку положительное влияние инокуляции распространяется и на корни растений, то после сбора урожая пожнивные остатки более эффективно действуют на последующую культуру севооборота.
Препарат, содержащий клубеньковые бактерии, готовят разными методами. Чаще всего используют торфяной нитрагин — ризоторфин. Он представляет собой стерилизованный у-облучением низинный торф, к которому добавлены необходимые для клубеньковых бактерий питательные вещества. Расфасованную массу выдерживают в термостате для размножения внесенных в нее бактерий. Иногда готовят торфяной препарат, не стерилизуя торф, но вносят в него большое количество клубеньковых бактерий.
Ризоторфин позволяет уменьшить объемы применения азотных удобрений; препарат разработан практически для всех бобовых, возделываемых в настоящий момент. Особенно ярко полезность ризоторфина проявляется при введении в культуру новых видов бобовых, многие из которых (например, козлятник) вообще не могут возделываться без наличия в почве соответствующих микроорганизмов. Агрономическая эффективность ризоторфина для бобовых культур в среднем составляет 10—30%, дополнительный сбор белка — 2—5 ц/га. При интродукции новых бобовых культур (люпин, люцерна, козлятник) эффективность бактеризации может составлять 50—100%, а сбор белка увеличивается в 2—3 раза.