Материал: микра от 104 группы
25. Ферменты микроорганизмов. Экзо- и эндоферменты микроорганизмов. Роль пермеаз (транслоказ) в жизнедеятельности микробной клетки.
Глубокое понимание процессов метаболизма микроорганизмов вряд ли возможно без предварительного знакомства с ролью и значением ферментов. Ферменты — биологические катализаторы. Они катализуют тысячи химических реакций, из которых слагается метаболизм организма. Известно уже около двух тысяч ферментов. По химической природе ферменты — глобулярные белки молекулярной массой от 10 000 до нескольких миллионов. Название ферменту во многих случаях дают по веществу, на которое он действует, с изменением окончания на «-аза». Например, целлюлаза катализует гидролиз целлюлозы до целлобиозы, уреаза — гидролиз мочевины (urea) до аммиака и СО2 и т. п. Однако чаще фермент получает наименование, которое указывает на природу катализуемой им химической реакции. Современная классификация ферментов также строится с учетом природы реакций, которыми они управляют. Согласно разработанной Комиссией по ферментам Международного биохимического союза классификации, выделяют шесть главных классов ферментов.
1. Оксидоредуктазы. Эти ферменты катализуют окислительно-восстановительные реакции, играют большую роль в процессах биологического получения энергии. К ним относятся дегидрогеназы (НАД, НАДФ, ФАД), цитохромы (b, с, с, а, а3 ), ферменты, участвующие в переносе водорода, электронов и др.
2. Трансферазы. Катализуют перенос отдельных радикалов, частей молекул или целых атомных группировок от одних соединений к другим. Например, ацетилтрансферазы переносят остатки ацетата —СН3СО, а также молекулы жирных кислот; фосфотрансферазы, или киназы, обусловливают перенос остатков фосфорной кислоты Н3РO3 2 - . Известны и другие трансферазы (аминотрансферазы, фосфорилазы и т. д.).
3. Гидролазы. Катализуют реакции расщепления и синтеза белков, жиров и полисахаридов с участием воды. К данному классу относят протеолитические ферменты (или пептидгидролазы), действующие на белки или пептиды; гидролазы глюкозидов, осуществляющие каталитическое расщепление углеводов и глюкозидов B-фруктофуранозидаза, a -глюкозидаза, а- и B-амилаза, B-галактозидаза и др.); эстеразы, катализующие расщепление и синтез сложных эфиров (липазы, фосфатазы).
4. Лиазы. Включают ферменты, катализующие отщепление от субстратов определенных химических групп с образованием двойных связей или присоединение отдельных групп радикалов к двойным связям. Так, пируватдекарбоксилаза катализует отщепление СО2 от пирувата:
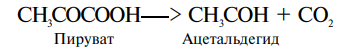
К лиазам относится также фермент альдолаза, расщепляющий шестиуглеродную молекулу фруктозо-1,6-бисфосфата на два трехуглеродных соединения.
5. Изомеразы. Участвуют в превращении органических соединений в их изомеры. При изомеризации происходит внутримолекулярное перемещение атомов, атомных группировок, различных радикалов и т. п. Изомеризации подвергаются углеводы и их производные, органические кислоты, аминокислоты и т. д. К данной группе относятся триозофосфатизомераза, глюкозофосфатизомераза и др. 6. Л и газы. Катализируют синтез сложных органических соединений из простых. Например, аспарагинсинтетаза управляет синтезом амида аспарагина из аспарагиновой кислоты и аммиака с обязательным участием АТФ, дающей энергию для этой реакции:
![]()
К группе лигаз относят карбоксилазы, катализующие присоединение СО2 к различным органическим кислотам. Например, фермент пируваткарбоксилаза катализует синтез оксалоацетата из пирувата и СО2 .
По связи с бактериальной клеткой различают:
Экзоферменты – ферменты не связанные со структурой протоплазмы, легко выделяющиеся в субстрат (гидролитические ферменты), растворимые в питательной среде и проходящие через бактериальные фильтры. Экзоферменты участвуют в процессе питания, расщепляя сложные высокомолекулярные соединения (крахмал, белки, клетчатку) подготавливая питательные вещества к усвоению микробной клеткой.
Эндоферменты – ферменты прочно связанные с бактериальной клеткой и действующие только внутриклеточно. Они осуществляют дальнейшее разложение питательных веществ и превращают их в составные части клетки. К таким ферментам относятся дегидрогеназы и оксидазы.
Пермеазы (permease)— ферментоподобные белки, участвующие в транспорте метаболитов, находятся в периплазме клетки. Эти белки участвуют в облегчённой диффузии веществ в клетку. Этим путём в клетку поступает ограниченное количество соединений.
Также существуют специфические пермеазы, локализованные в цитоплазматической мембране. Которые обеспечивают активный транспорт внутрь клетки. Данный процесс требует энергетических затрат и происходит даже в тех случаях, когда концентрация необходимых веществ в среде ниже, чем в микробной клетке. При этом каждая пермеаза переносит в клетку только определённую аминокислоту или другое соединение.
В процессе переноса может произойти химическая модификация вещества — транслокация химических групп, например фосфорилирование углеводов при участии соответствующих ферментов.
26. Превращение микроорганизмами соединений азота. Значение этих процессов в природе и с/х.
От азотного питания растений во многом зависит величина урожая сельскохозяйственных культур. Большинству растений недоступен газообразный азот, в огромном количестве находящийся в воздухе, а из разнообразных азотных соединений, встречающихся в почве, они могут усваивать только минеральные. Поэтому столь важен вопрос о превращениях соединений азота в почве под воздействием микроорганизмов. Превращения азота и содержащих этот элемент соединений в почве довольно сложны, но в них можно выделить основные направления, определяющие круговорот азота в природе:
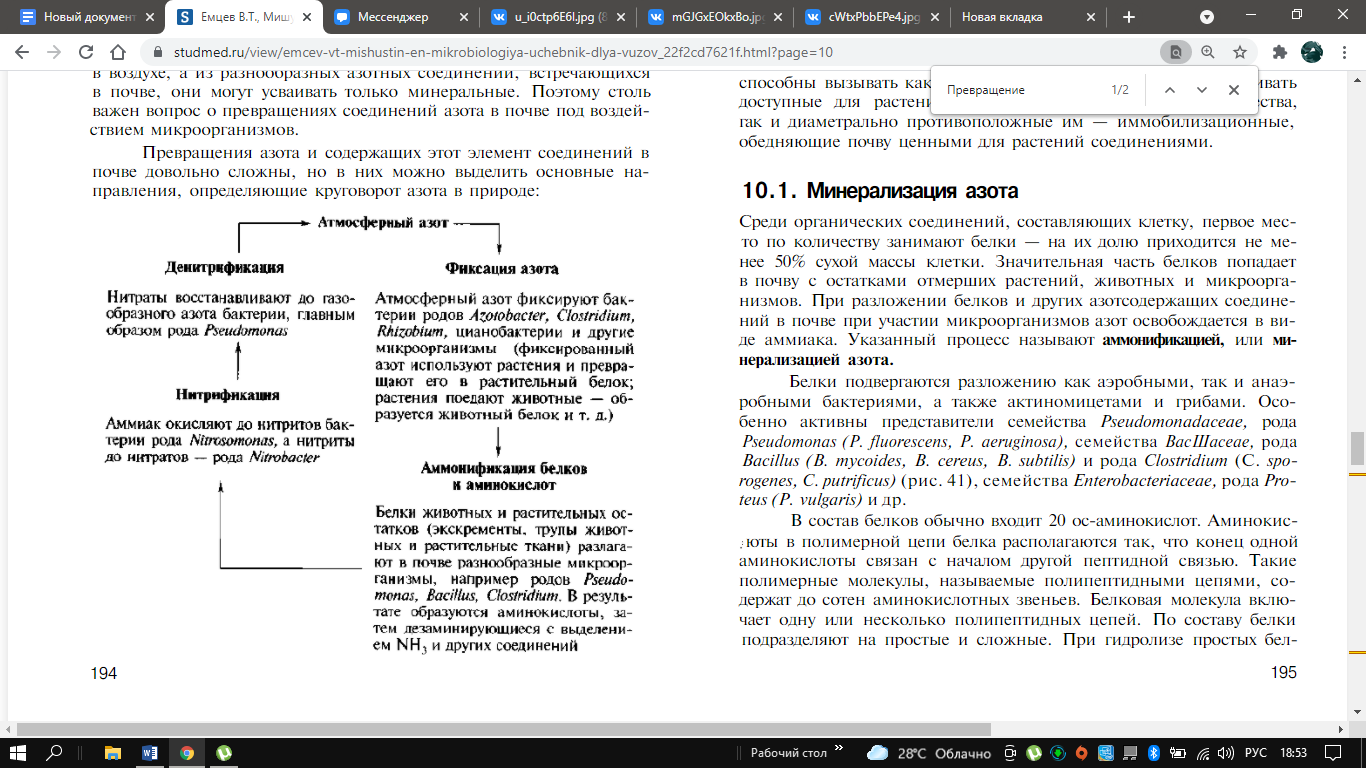 Некоторую
часть атмосферного азота связывают
свободноживущие или находящиеся в
симбиозе с растениями микроорганизмы.
Данный процесс обогащает азотом и почву,
и растения. Органические азотсодержащие
соединения в тканях растений и животных,
попадая в почву, подвергаются минерализации
до аммонийных соединений. Часть
растительных остатков трансформируется
в темноокрашенное, содержащее азот
вещество, — гумус. Аммонийная форма
азота подвергается в почве окислению
нитрифицирующими бактериями с образованием
солей азотной кислоты. При определенных
условиях нитраты могут восстанавливаться
до молекулярного азота и улетучиваются
из почвы. Значительное количество
азотсодержащих соединений микроорганизмы
ассимилируют, а азот в органических
формах практически недоступен растениям.
Приведенные примеры показывают, что
микроорганизмы способны вызывать как
мобилизационные процессы и накапливать
доступные для растений минеральные
азотсодержащие вещества, так и диаметрально
противоположные им — иммобилизационные,
обедняющие почву ценными для растений
соединениями.
Некоторую
часть атмосферного азота связывают
свободноживущие или находящиеся в
симбиозе с растениями микроорганизмы.
Данный процесс обогащает азотом и почву,
и растения. Органические азотсодержащие
соединения в тканях растений и животных,
попадая в почву, подвергаются минерализации
до аммонийных соединений. Часть
растительных остатков трансформируется
в темноокрашенное, содержащее азот
вещество, — гумус. Аммонийная форма
азота подвергается в почве окислению
нитрифицирующими бактериями с образованием
солей азотной кислоты. При определенных
условиях нитраты могут восстанавливаться
до молекулярного азота и улетучиваются
из почвы. Значительное количество
азотсодержащих соединений микроорганизмы
ассимилируют, а азот в органических
формах практически недоступен растениям.
Приведенные примеры показывают, что
микроорганизмы способны вызывать как
мобилизационные процессы и накапливать
доступные для растений минеральные
азотсодержащие вещества, так и диаметрально
противоположные им — иммобилизационные,
обедняющие почву ценными для растений
соединениями.
27. Структура микробных сообществ почв различных типов и факторы, определяющие её формирование.
Численный состав микроскопических существ почв отличается большой динамичностью. Даже за относительно короткие промежутки времени число микроорганизмов в почве может значительно меняться. Это следствие динамики температуры и влажности почвы, состояния растительного покрова и г. д. Почти во всех почвах наблюдается большая или меньшая активизация деятельности микроорганизмов весной.Кроме сезонных изменений, в численности почвенной микрофлоры отмечаются и кратковременные флуктуации. О причине последних существуют разные предположения. Некоторые исследователи допускают, что число бактерий может резко снижаться вследствие уничтожения их фагами или простейшими. Предполагают также накопление каких-то токсичных веществ в почве (этилена, окиси этилена и др.), временно подавляющих развитие определенных групп микроорганизмов.Однако скорее всего флуктуации определяются неравномерным распределением микроорганизмов в почве.
Так же, по мере перехода от более холодного северного климата к южному микронассленис почв возрастает; во многих южных почвах и микробиологические процессы протекают более энергично.
Наиболее изучена сапротрофная. или зимогенная (от греч. туте — закваска), группировка микроорганизмов у различных почв, т. е. микроорганизмы, разлагающие в основном легко доступные органические соединения. Обычно сапротрофов учитывают методом посева на твердые, а иногда и в жидкие питательные среды, содержащие те или иные органические вещества. Наиболее часто используют мясо-пеп- тонный агар и крахмало-аммиачный агар, на которых хорошо выявляются бактерии и, в частности, актином и цеты.
усредненные данные о численности и соотношение основных групп сапротрофных микроорганизмов в верхних слоях различных почв
Зоны |
Почвы |
Состояние ПОЧВ |
Общее число микроорганизмов, тыс. на 1 г почвы |
Бактерии |
Споры (из числа бактерий) |
Актиномицеты |
Грибы |
% |
|||||||
Тундра и тайга |
Ту ндрово-глссвыс и глеево-подзол истые |
Целинные |
2140 |
95,6 |
0,7 |
1*4 |
3,0 |
|
|
Окультуренные |
4870 |
98,0 |
0,6 |
1,6 |
0,4 |
Лесо-луговая |
Подзолы и дерново- подзол истые |
Целинные |
1080 |
89,3 |
12,0 |
8,1 |
2,6 |
|
|
Окультуренные |
2620 |
70,7 |
14,9 |
28,2 |
1,1 |
Луговая степь и степь |
Черноземы |
Целинные |
3630 |
63,8 |
21,4 |
35,4 |
0,8 |
|
|
Окультуренные |
4530 |
64,4 |
24,5 |
35,1 |
0,5 |
Сухая степь |
Каштановые |
Целинные |
3480 |
64,8 |
19,3 |
34,7 |
0,5 |
|
|
Окультуренные |
6660 |
67,6 |
23,0 |
32,0 |
0,4 |
Пустынная степь и пустыня |
Бурые и сероземы |
Целинные |
4490 |
63,4 |
17,7 |
36,1 |
0,5 |
|
|
Окультуренные |
7380 |
66,1 |
19,8 |
33,6 |
0,3 |
Как видно из приведенных данных, в составе зимогенной микрофлоры богато представлены бактерии, особенно неспорообразующие формы. Однако количество их в разных почвах неодинаково. Так, можно считать доказанным, что гнилостные бактерии Pseudomonas fluorescens, являющиеся пионерами освоения органических растительных остатков, более богато представлены в почвах севера, где медленно идет минерализация. В почвах юга они обнаруживаются в значительном числе лишь в течение краткого времени после внесения растительных остатков.
Представители рода Arthrobacter, по ряду признаков родственные актиномицстам, в большем числе встречаются в почвах южной зоны. Они характерны для более поздних стадий распада органического вещества и предпочитают нейтральную среду. В почвах севера очень часто присутствует значительное количество кори- небактерий.
Из неспорообразующих азотфиксирующих бактерий виды рода Beijerinkia распространены только в кислых субтропических почвах (латеритах и желтоземах). Представители рода Enterobacter в большом количестве встречаются в лесных почвах средней полосы, а рода Spirillum — в южной зоне.
! Основательнее изучены группировки спорообразующих бактерий, развитие которых связано с присутствием в почве более переработанного органического вещества.
Каждому типу почв свойствен характерный набор преобладающих видов бацилл. Другие виды здесь могут быть, но в очень малом количестве.
Доп инфа. https://studme.org/287642/ekologiya/struktura_mikrobnyh_soobschestv_pochv_raznyh_tipov
28. Питание микроорганизмов. Способы питания и поступления питательных веществ в клетку. Источники отдельных питательных элементов (углерода, азота и др)
Питание бактерий – это процесс поглощения и усвоения бактериальной клеткой пластического материала и энергии в результате преобразовательных реакций[4].
Питание является неотъемлемой функцией каждого живого организма. В процессе питания организм получает вещества, идущие на синтез клеточных структур и служащие источником энергии для всех процессов жизнедеятельности. Для питания микроорганизмов необходимы те же элементы, что и для животных, и растений. Первоочередные элементы питания – углерод, азот, кислород, водород, являющиеся основой всех органических веществ, которые входят в состав живой клетки как прокариоритеческих так и эукариоэтических организмов[5].
Типы питания бактерий чрезвычайно разнообразны. Различаются они в зависимости от способа поступления питательных веществ бактериальной клетки, источников углерода и азота, способа получения энергии, природы доноров электронов
Способы поступления питательных веществ
Источники углерода
Источники энергии
Природа доноров электронов
Источники углерода, энергии и доноров электронов
Источники азота
Способы поступления питательных веществ
По способам поступления питательных веществ бактерии подразделяются на:
голофиты (греч. holos – полноценный и греч. phyticos – относящийся к растениям) – бактерии неспособные выделять в окружающую среду ферменты, расщепляющие субстраты, потребляют вещества только в растворенном, молекулярном виде;
голозои (греч. holos – полноценный и греч. zoikos – относящийся к животным) – бактерии, обладающие комплексом ферментов, обеспечивающие внешнее питание – расщепление субстратов до молекул вне бактериальной клетки, после чего молекулы питательных веществ транспортируются внутрь бактерии.
Источники углерода
По источникам углерода различают:
автотрофы (греч. autos– сам, trophe – пища) – бактерии, использующие в качестве источника углерода углекислый газ (CO2), из которого осуществляют синтез всех углеродосодержащих веществ;
гетеротрофы (греч.geteros– другой, trophe– пища) – бактерии, использующие в качестве источника углерода различные органические вещества в молекулярной форме (многоатомные спирты, углеводы, жирные кислоты, аминокислоты)[4].
Наибольшая степень гетеротрофности отмечается у прокариот, живущих только внутри других живых клеток, в частности хламидий и риккетсий.
Источники энергии
В зависимости от используемых источников энергии бактерии подразделяют на два типа:
фототрофы – бактерии способные использовать солнечную энергию;
хемотрофы – бактерии, получающие энергию при окислительно-восстановительных реакциях[4].
Природа доноров электронов
По природе доноров электронов бактерии делят на:
литотрофы (греч. litos – камень) – бактерии, использующие в качестве доноров электронов неорганические вещества: водород (Н2), сероводород (Н2S), аммиак (NH3), серу (S), углекислый газ(CО2), ионы железа (Fe2+) и многие другие;
органотрофы – бактерии, использующие в качестве донора электроноворганические соединения (углеводы, аминокислоты)[4].
В зависимости от источника энергии и природы донора электроноввозможно четыре основных типа энергетического метаболизма: хемолитотрофия, хемоорганотрофия, фотолитотрофия, фотоорганотрофия. Таки образом, бактерии разделяют на: