Материал: метаболизм аминокислот
МЕТАБОЛИЗМ АМИНОКИСЛОТ
Катаболизмаминокислот ициклмочевины |
|
|
|
|||||||
Большую частьметаболическойэнергии,вырабатываемой втканях, поставляют |
|
|||||||||
процессыокисленияуглеводовитриглицеридов,увзрослогомужчины до90% всей |
||||||||||
потребностивэнергиипокрываетсяизэтихдвухисточников. Остальную энергию |
||||||||||
даетокисление аминокислот. Приокисленииаминокислот,кроме этого, |
|
|||||||||
|
|
|
|
начина тсясудаления |
α |
-аминогруппы |
|
|||
синтезируютсямногие биологическиактивные соединения. |
|
. |
||||||||
Катаболизм большинствааминокислот |
|
|
|
|
|
|
||||
трансаминирования |
|
дезаминирования |
|
|
|
|
|
|||
Аминокислотатеряетаминогруппув результатедвухтипов реакций: |
|
|||||||||
|
|
и |
|
|
|
. |
|
|
|
|
Первыйэтапзаключаетсявобратимом переносе NH2-группы саминокислотына |
|
|||||||||
|
трансаминирование |
|
|
|
|
|
|
|||
кетокислотусобразованиемновойаминокислотыиновойкетокислоты –этот |
|
|||||||||
переносназывается |
|
|
|
|
|
|
α-ке оглутаровуюкислоту, |
|
||
|
|
|
|
и егомеханизмдовольносложен. |
|
|||||
Ворганизмеаминогруппаобычно переноситсяна |
|
|
|
|
||||||
которая превращаетсявглутамат (глутаминоваякислота). |
|
|
|
|||||||
Реакциикатализируютферменты ,коферментомкоторых являетсяпиридоксальфосфат —производноеам нотрансферазывитаминаВ6 (пиридоксина). Пиридоксальфосфатслужит переносчиком аминогрупп. Вступать вреакции трансаминированиямогутпочти все аминокислоты,заисключениемлизина, треонинаипролина. Аминотрансферазы —классическийпримерферментов, катализирующихреакции,протекающие по механизмутипа«пинг-понг».Втаких реакцияхпервый продукт долженуйтиизактивногоцентраферментадотого,как второйсубстратсможеткнемуприсоединиться.
Аминотрансферазы обладаютсубстратной специфичностью кразным аминокислотам.Втканяхчеловека обнаруженоболее 10разныхаминотрансфераз.
|
|
|
АЛТ), |
|
(ACT). |
Наиболее распространённымиферментами( |
вбольшинстве тканеймлекопитающих |
||||
являютсяаланинаминотрансфераза |
|
α |
|
||
|
иаспартатаминотрансфераза |
|
|||
АЛТ катализируетреакцию трансаминированиямеждуаланиноми -кетоглута- |
|||||
L-аланин+α-кетоглутарат |
↔ |
пируват+ L-глутамат |
|
||
ратом: |
|
|
|
|
|
ACT катализируетреакцию трансаминированиямеждуаспартатоми -кетоглута- ратоманалогичнопредыдущей: α
L-аспартат+α-кетоглутарат ↔ оксалоацетат+ L- глутамат
Наибольшее количествоАЛТ иACT сосредоточеновпечениимиокарде,а содержание вкровиоченьнизкое, поэтому ихиспользуютдля диагностикиэтих органов.При поврежденииклетоксоответствующегооргана ферментывыходят в
кровь.Увеличение их активностиуказываетнастепеньиглубинуразрушения органов(инфарктмиокарда,гепатитит.д.).
В клетках сердечноймышцы количество АСТпревышает АЛТ, ав печенинаоборот. Поэтому особенно информативно одновременное измерение активностиобоих ферментов всыворотке крови. Соотношение активностиАСТ/АЛТназывают “коэффициентдеРитиса”,который в нормеравен 1.33+-0.42. при инфаркте миокардаактивность АСТ в кровиувеличиваетсяв 8-10 раз, поэтому коэффициент ре Ритиса сильно возрастает. При гепатитах же этот показатель снижается до
0,6.
трансаминированиясостоитвтомчтоаминогруппыот многихразных |
|||||||
аминокислотсобираются |
|
|
|
– ввиде глутаминовой кислоты. |
|||
С ысл |
|
|
|
|
|
|
|
Последнийиспользуетсядлядезаминированияаминокислот,содержащихся в |
|||||||
|
одной форме |
|
|
|
|
||
митохондриях. Этоочень |
важно |
,таккак |
то ькоглутамат |
втканяхмлекопитающих |
|||
наиболее быстро можетподвергаться окислительному дезаминированию. |
|||||||
|
|
реакцияотщепления |
-аминогруппыот |
||||
аминокислоты,в результате чегообразуетсясоответствующая. |
-кетокислота |
||||||
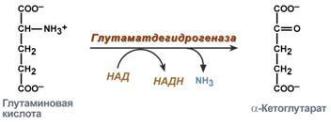
Дезаминированиеглутамата- |
|
|
|
α |
|
|
|
(безазотистыйостаток)ивыделяетсямолекулааммиака |
|
α |
|||||
|
|
|
|
|
|
|
|
глутамат + NAD+ + H2O→ α-кетоглутароваякислота + NADH + H+ + NH4+
Окислительное дезаминирование глутаматакатализируетсяферментом |
|
|||
Глутаматдегидрогеназа . |
|
|
|
|
играетважную роль,таккакявляется регуляторным |
||||
ютаматдегидроген за |
|
|
|
|
ферментомаминокислотногообмена. Алостерические ингибиторы |
|
|||
глутаматдегидрогеназы(АТФ,ГТФ,NADH)вызывают диссоциацию ферментаи |
||||
потерю глутаматдегидрогеназнойактивности.Высокие концентрацииАДФ |
||||
активируют фермент. Такимобразом,низкийэнергетическийуровеньвклетках |
||||
стимулируетразрушение аминокислотиобразование -кетоглутарата, |
|
|||
поступающеговЦТКкак энергетическийсубстрат.Реакцииокислительного |
||||
|
|
α |
|
|
дезаминированияидутвклеткахвсехорганов,(кроме мышц),но особенно |
|
|||
интенсивноонипротекаютвклеткахпеченимитохондриальном. |
матриксе |
. |
||
Глутаматдегидрогеназанаходитсятольков |
|
|||
Катаболизмаминокислотв тканяхпроисходитпостояннойскоростью —100г/сут. |
||||
Приэтомв результате дезаминированияаминокислотосвобождаетсябольшое |
||||
количествоаммиака, которыйтоксичен дляЦНС,поэтомувклеткахактивно |
||||
происходитсвязывание аммиакасобразованиемнетоксичныхсоединений.Одним |
||||
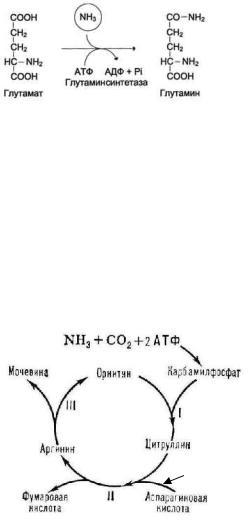
изтакихсоединенийявляется |
|
,которыйобразуетсяпод действием |
||
глутамин-синтетазы: |
глутамин |
|
|
|
|
|
|
|
|
Глутаминявляетсянейтральным,нетоксическим веществом, он доставляетсякровью впечень,где образуютсяконечные продукты азотногообменаи выводятсяизорганизма.
Выведен еаминногоазотаизорганизма: |
|
|
|
|
||||||||||
Ворганизме человекаимлекопитающих аминныйазотпревращаетсяв |
||||||||||||||
нетоксичное, хорошо растворимое соединение — |
|
|
,ввиде которогои |
|||||||||||
выводится изорганизма. |
|
|
|
|
|
|
мочевину |
|
|
|
||||
Мочевинаобразуетсяв |
|
|
мочевина |
|
,или |
|
|
|
|
|
,который |
|||
происходитв |
|
, затем |
|
|
доставляетсякровью в почкиивыводитсяс |
|||||||||
|
|
|
цикле |
|
ны |
|
орни ин |
омцикле |
|
|||||
мочой.Синтезмочевины -циклический процесссостоитизпяти реакций, |
||||||||||||||
|
печени |
|
|
|
|
|
|
|
|
|
|
|
|
|
катализируемый пятью отдельнымиферментами |
|
|
|
|
|
|||||||||
|
|
|
|
|
|
Изанализа процессасинтезамочевины |
||||||||
|
|
|
|
|
|
следует: |
|
|
|
|
|
|
||
|
|
|
|
|
|
1.включение азотапроисходитв двухточках. |
||||||||
|
|
|
|
|
|
Одинизатомовазотапоступаетв форме NH3 и |
||||||||
|
|
|
|
|
|
являетсяпродуктомдезаминирования |
||||||||
|
|
|
|
|
|
аминокислот,адругой включаетсявсоставе |
||||||||
|
|
|
|
|
|
аспартата. Этот второйазотможетпоступатьв |
||||||||
|
|
|
|
АТФ |
аспартатизлюбойаминокислоты путем |
|||||||||
|
|
|
|
|
|
трансаминированиясоксалоацетатом. |
||||||||
|
|
|
|
|
|
Следовательно,атомыазотавмочевине имеют |
||||||||
|
|
|
|
|
|
разное происхождение; |
|
|
||||||
2.орнитиновыйцикл связансцитратнымциклом,таккакоксалоацетат, |
||||||||||||||
необходимый длятрансаминирования,образуетсяизфумаратав реакциях |
||||||||||||||
цитратногоцикла; |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
3. процессэндергонический,требующий 3 молекулАТР(4фосфатныхгрупп) для |
||||||||||||||
синтезаодноймолекулымочевины. |
|
|
|
|
|
|
|
|
|
|||||
Принедостаточнойактивностиферментов орнитиновогоциклавозникают |
||||||||||||||
гипераммониемии - патологические состояниясопровождающиесяповышением |
||||||||||||||
концентрацииаммиакавкрови. |
|
|
|
|
|
|
|
|
|
|||||
После удаленияаминогруппв реакцияхтрансаминированияс α-кетоглутаратом |
||||||||||||||
углеродные скелетыаминокислотподвергаютсяокислительному расщеплению и |
||||||||||||||
превращаютсявсоединения,способные включатьсявцикллимонной кислотыи |
||||||||||||||
окислятьсявнем.Существует20 разныхпутейрасщепленияуглеродногоскелета |
||||||||||||||
для20различныхаминокислот. из20аминокислотобразуется 5 продуктов цикла |
||||||||||||||
Кребса.Они включаются вциклипродолжают окисление доСО2 |
иН2О. |
|||||||||||||
Декарбоксилированиеаминокислоткарбоксильнойметаболизм биогенных аминов
Врезультате отщепления - группыаминокислот образуются амины. Реакциякатализируетсяα декарбоксилазами,коферментом которых является

фосфопиридоксаль. Продукты декарбоксилированияобладаютвысокой биологическойактивностью исэтимсвязаноихназвание -биогенные амины.
образуется изгистидинавтучныхклетках.Выделяется вответна присутствиеГистамин аллергена.Кроме того, являетсясильнымсосудорасширяющим фактором,вызываетсокращение гладкоймускулатуры,вклетках слизистой желудкастимулирует секрецию солянойкислоты.
(ГАМК)образуетсяизглутаматавтканиголовного мозгаγ-аминомасл,выполняетнаяфункциикислотатормозногонейромедиатора.
образуетсяизтриптофанавнейронахгипоталамуса. Функционируеткак возможныйСерото ин нейромедиаторвозбуждающегохарактера.
образуется изтирозинавпочках, надпочечниках,синаптическихганглиях, нервахДоф мин.Являетсямедиаторомингибирующеготипа,функционируетвчерной субстанцииверхнегоотделастволамозга. Вдругихклеткахявляется предшественникомнорадреналинаиадреналина.
образуетсяврезультате гидроксилированиядофаминавклетках нервнойНорадреналинткани,мозговомвеществе надпочечников.Функционируеткакмедиатор в синаптическойпередаче нервныхимпульсов.
-продукт метилированиянорадреналинавклеткахмозговоговещества надпочечниковАдреналин .Выполняетфункциигормона.
Инактивациябиогенныхаминовпроисходитпутемих дезаминированияи окисления.Реакцию катализируетFAD-зависимаямоноаминооксидаза(МАО). Моноаминооксидазаможетбытьточкойвоздействиянекоторых лекарств, ингибирующихилиактивирующихэтотфермент,таккакизменение концентрации биогенныхаминовявляетсяпричиной ряда патологическихсостояний.Например, при паркинсонизме наблюдаетсяуменьшение количествадофамина,иоднимиз способовлеченияявляетсяснижение скоростиинактивациидофаминапод влияниемвеществ-ингибиторовМАО.
Биосинтез аминокислот
Ватмосфере элементарныйазот(Ν2)присутствует практическив неограниченном количестве.Прежде чемпоступитьвкруговоротазота,он должен быть восстановлендо NH3 ивключен ваминокислоты. Фиксировать атмосферныйазот способнылишьнемногие виды бактерий и синезеленыхводорослей,которыеиз атмосферногоазота образуютаммиак -NH3 ,этот процессноситназвание
. Бактерии-азотофиксаторы находятся впочве свободноили живут вазотофиксациисимбиозе срастениями.Особоважное значение имеетсимбиозмежду бактериями ибобовымирастениями такими,какклевер,бобыилигорох(этирастенияочень питательныблагодарявысокому, содержанию белка).Затемаммиакиспользуется либонепосредственно,либопочвеннымибактериямипревращаетсяврастворимые соединения(нитриты инитраты) впроцессе нитрификации.
В симбиозес бобовымибактерии живут в корневыхклубочках внутри растительных клеток, так называемые бактероидах. С одной стороны, растение снабжает бактериоды питательнымивеществами, ас другой, извлекает пользу от фиксированного азота, которыйпоставляет симбионт. Фиксирующим
N2 ферментом бактерий является нитрогеназа.Онасостоит из двух компонентов:Fe-белкаиFeMo-белка. Fe-белок, содержащий[FeS4]-центр, служит окислительно-восстановительной системой, которая принимает электроны отферредоксина ипередает их во второйкомпонент,FeMo-белок. Этот молибденсодержащий белок переносит электроны на N2 итаким образом через различные промежуточные стадии продуцирует NH3. Часть восстановительных эквивалентов переносится в побочнойреакциина H+. Поэтому наряду с NH3всегда образуется водород.
Втовремякак растениямлекопитающиемикроорганизмы могутвполне синтезироватьвсе |
|
|||
аминокислоты, |
входе эволюцииутратилиспособностьксинтезу |
|||
|
|
незаменимымиаминокис |
ами |
|
примерно половиныиз20аминокислот. Остальные аминокислоты должны |
|
. |
||
поступатьворганизмспищей,иназываются |
|
|
||
Заменимыеам |
окислоты (аланин,аспарагиноваяиглутаминоваякислотыиих |
|||
амиды,аспарагиниглутамин)образуютсяв результате трансаминированияиз промежуточныхметаболитов —образующихсяв процессахгликолизаицикла лимоннойкислоты(циклаКребса),апредставителисериновогосемейства(серин, глициницистеин)самиявляютсяестественнымиметаболитами организма животных.