Материал: Лекция_53-54
Взаимопревращения в щелочном растворе между альдозами, эпимерами по с-2, называют эпимеризацией. Получение гликаровых кислот
П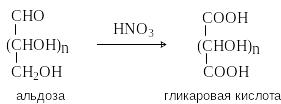 ри
действии разбавленной азотной кислоты
окисляется оба конца углеродного скелета
альдоз и образуются гликаровые
кислоты.
ри
действии разбавленной азотной кислоты
окисляется оба конца углеродного скелета
альдоз и образуются гликаровые
кислоты.
При образовании гликаровых кислот, как и в случае глицитов, происходит «уравнивание» функциональных групп на концах цепи и из некоторых альдоз образуются мезо-соединения.
Окисление кетоз азотной кислотой протекает с расщеплением С-С связей.
Окисление виц-диольной группировки
Моносахариды содержат виц-диольную группировку, которая претерпевает окислительное расщепление под действием иодной кислоты или тетраацетата свинца.
HOCH2(CHOH)nCHO + (n+1) HIO4 (n+1) HCOOH + HCHO + (n+1) HIO3
Р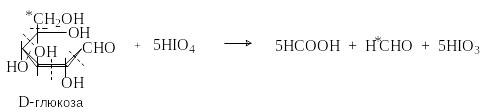 еакция
протекает количественно. По составу
продуктов и количеству израсходованной
иодной кислоты получают ценную информацию
о строение моносахаридов и их производных.
еакция
протекает количественно. По составу
продуктов и количеству израсходованной
иодной кислоты получают ценную информацию
о строение моносахаридов и их производных.
Получение гликозидов
Циклические формы моносахаридов содержат несколько групп ОН, одна из которых – гликозидный (полуацетальный) гидроксил, отличается повышенной склонностью к реакциям нуклеофильного замещения.
И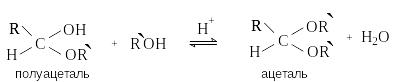 звестно,
что в присутствии кислотных катализаторов
полуацетали реагируют со спиртами. При
этом происходит нуклеофильное замещение
полуацетального гидроксила и образуются
полные ацетали.
звестно,
что в присутствии кислотных катализаторов
полуацетали реагируют со спиртами. При
этом происходит нуклеофильное замещение
полуацетального гидроксила и образуются
полные ацетали.
А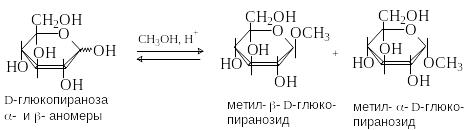 налогично
реагирует гликозидный гидроксил в
альдозах и кетозах. Спиртовые ОН группы
при этом не затрагиваются. Продукты
замещения гликозидного гидроксила
называют гликозидами
(гликопиранозидами
или гликофуранозидами
в зависимости
от размера цикла). Например, при пропускании
через раствор D-глюкозы в метаноле
газообразного HCl образуется смесь
метилглюкозидов, соответствующих разным
таутомерным формам D-глюкозы (двум
пиранозным и двум фуранозным). В условиях
термодинамического контроля в реакционной
смеси преобладают более стабильные
метилпиранозиды.
налогично
реагирует гликозидный гидроксил в
альдозах и кетозах. Спиртовые ОН группы
при этом не затрагиваются. Продукты
замещения гликозидного гидроксила
называют гликозидами
(гликопиранозидами
или гликофуранозидами
в зависимости
от размера цикла). Например, при пропускании
через раствор D-глюкозы в метаноле
газообразного HCl образуется смесь
метилглюкозидов, соответствующих разным
таутомерным формам D-глюкозы (двум
пиранозным и двум фуранозным). В условиях
термодинамического контроля в реакционной
смеси преобладают более стабильные
метилпиранозиды.
Метил--D-глюкопиранозид большей термодинамической стабильностью, чем -аномер (аномерный эффект) и поэтому образуется в большем количестве. Гликозиды существуют только в циклической форме, поэтому - и -аномеры гликозидов не могут спонтанно переходить друг в друга в результате таутомерных превращений. Гликозиды не имеют свободной альдегидной группы и являются невосстанавливающими сахарами.
К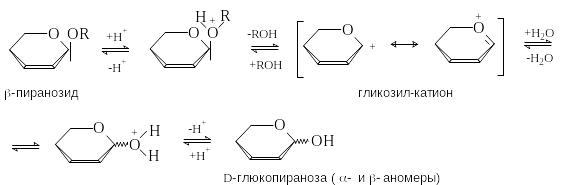 ак
полные ацетали гликозиды гидролизуются
в условиях кислотного катализа и
устойчивы в разбавленных растворах
щелочей. Механизм кислотного гидролиза
включает протонирование гликозидного
кислорода, расщепление гликозидной С-О
связи с образованием гликозил-катиона,
который затем атакуется молекулой воды.
ак
полные ацетали гликозиды гидролизуются
в условиях кислотного катализа и
устойчивы в разбавленных растворах
щелочей. Механизм кислотного гидролиза
включает протонирование гликозидного
кислорода, расщепление гликозидной С-О
связи с образованием гликозил-катиона,
который затем атакуется молекулой воды.
Расщепление гликозидной связи важно с биологической точки зрения, поскольку многие природные соединения являются гликозидами. Широко используется ферментативный гидролиз гликозидов, преимущество которого заключается в его специфичности. Определенные ферменты гидролизуют только - или только -гликозидные связи., что может быть использовано для установления конфигурации гликозидной связи.
М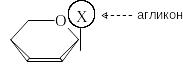 олекулу
гликозида рассматривают как состоящую
из двух частей – сахарной части и
агликона:
олекулу
гликозида рассматривают как состоящую
из двух частей – сахарной части и
агликона:
В качестве агликона в природных гликозидах могут выступать спирты, фенолы, стероиды, сами моносахариды. Перечисленные агликоны связаны с сахарной частью через атом кислорода, поэтому такие гликозиды называют О-гликозидами. В природе широко распространены N-гликозиды, в которых агликонами являются азотистые основания. К ним относятся нуклеозиды – структурные единицы нуклеиновых кислот.
Получение простых эфиров
М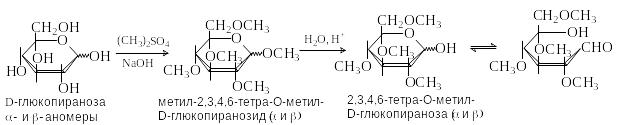 етиловые
эфиры по спиртовым группам ОН получают
действием на моносахариды диметилсульфата
в водном растворе щелочи или метилиодида
в присутствии оксида серебра. Эти методы
являются модификациями синтеза
Вильямсона. При этом в реакцию вступает
и гликозидный гидроксил. Простые эфиры
по спиртовым группа устойчивы к гидролизу,
в то время как гликозидная связь легко
расщепляется в кислой среде.
етиловые
эфиры по спиртовым группам ОН получают
действием на моносахариды диметилсульфата
в водном растворе щелочи или метилиодида
в присутствии оксида серебра. Эти методы
являются модификациями синтеза
Вильямсона. При этом в реакцию вступает
и гликозидный гидроксил. Простые эфиры
по спиртовым группа устойчивы к гидролизу,
в то время как гликозидная связь легко
расщепляется в кислой среде.
П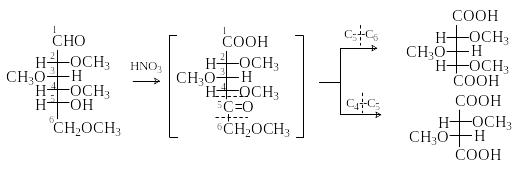 риведенная
последовательность реакций (метилирование,
затем гидролиз) используется для
определения размера цикла в моносахаридах.
Неметилированной остается группа ОН,
которая участвовала в образовании
циклического полуацеталя. Окисление
образовавшейся тетраметил-D-глюкозы
азотной кислотой в жестких условиях
дает сначала кетокислоту, а затем
триметоксиглутаровую и диметоксиянтарную
кислоты.
риведенная
последовательность реакций (метилирование,
затем гидролиз) используется для
определения размера цикла в моносахаридах.
Неметилированной остается группа ОН,
которая участвовала в образовании
циклического полуацеталя. Окисление
образовавшейся тетраметил-D-глюкозы
азотной кислотой в жестких условиях
дает сначала кетокислоту, а затем
триметоксиглутаровую и диметоксиянтарную
кислоты.
Состав продуктов окисления указывает на то, что кетогруппа, а, следовательно, и свободная гидроксигруппа находились в положении 5. Это означает, что цикл был пиранозным.
Получение сложных эфиров
Гидроксильные группы моносахаридов легко этерифицируются действием ангидриридов и хлорангидридов карбоновых кислот. Чаще всего используют ацетилирование уксусным ангидридом в присутствии кислотного (H2SO4, ZnCl2) или основного (пиридин, СH3COONa) катализатора.
С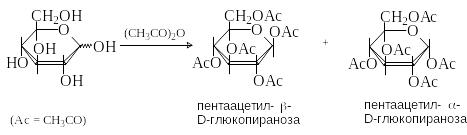 оотношение
-
и -аномеров
зависит от условий проведения реакции.
В условиях термодинамического контроля
(высокая температура, кислотный
катализатор) преобладает более стабильный
-аномер
(аномерный эффект). В условиях кинетического
контроля (температура ниже 00С,
основной катализатор) преимущественно
образуется -аномер,
поскольку экваториальная группа ОН
ацилируется с большей скоростью, чем
аксиальная.
оотношение
-
и -аномеров
зависит от условий проведения реакции.
В условиях термодинамического контроля
(высокая температура, кислотный
катализатор) преобладает более стабильный
-аномер
(аномерный эффект). В условиях кинетического
контроля (температура ниже 00С,
основной катализатор) преимущественно
образуется -аномер,
поскольку экваториальная группа ОН
ацилируется с большей скоростью, чем
аксиальная.
Д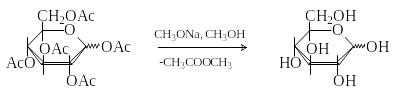 ля
удаления ацетильных групп используют
переэтерификацию действием метилата
натрия в метаноле.
ля
удаления ацетильных групп используют
переэтерификацию действием метилата
натрия в метаноле.
Получение производных по карбонильной группе. Образование озазонов
М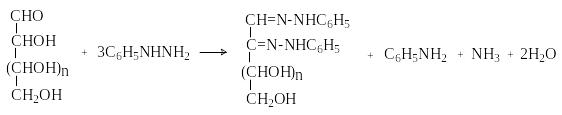 оносахариды
вступают во многие реакции нуклеофильного
присоединения по карбонильной группе,
характерные для альдегидов и кетонов:
присоединяют HCN,
NH2OH,
фенилгидразин. При действии избытка
фенилгидразина образуются озазоны.
оносахариды
вступают во многие реакции нуклеофильного
присоединения по карбонильной группе,
характерные для альдегидов и кетонов:
присоединяют HCN,
NH2OH,
фенилгидразин. При действии избытка
фенилгидразина образуются озазоны.
Эпимеры по С-2, например, D-глюкоза и D-манноза, дают один и тот же озазон, что используется для установления стереохимической конфигурации моносахаридов. Кетозы также образуют озазоны. D-фруктоза дает такой же озазон, что и D-глюкоза.
Озазоны – желтые кристаллические вещества, используются для идентификации сахаров.
Синтез моносахаридов
Полный синтез моносахаридов – очень сложная задача, так как при его осуществлении возникает необходимость разделения оптических изомеров. Обычно доступные из природных источников моносахариды используют для получения менее доступных сахаров. Для этого используют методы деградации и наращивания цепи, изменение конфигурации хиральных центров. Методы наращивания и деградации цепи, позволяющие производить переходы триозатетрозапентозагексоза и обратные превращения, имеют большое значение для установления конфигурации моносахаридов.
Наращивание цепи по методу Килиани-Фишера
М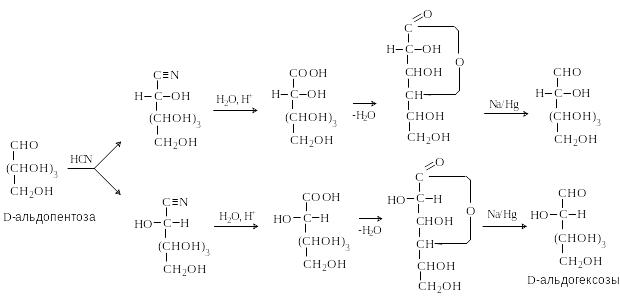 етод
включает присоединение HCN
по карбонильной группе, гидролиз
циангидринов до гликоновых кислот,
лактонизацию образующейся кислоты,
восстановление лактонов до альдоз. В
результате образуются две альдозы –
эпимеры по C-2,
так как на стадии образования циангидрина
появляется новый хиральный центр и
образуются два диастереомера.
етод
включает присоединение HCN
по карбонильной группе, гидролиз
циангидринов до гликоновых кислот,
лактонизацию образующейся кислоты,
восстановление лактонов до альдоз. В
результате образуются две альдозы –
эпимеры по C-2,
так как на стадии образования циангидрина
появляется новый хиральный центр и
образуются два диастереомера.
Деградация по Волю
О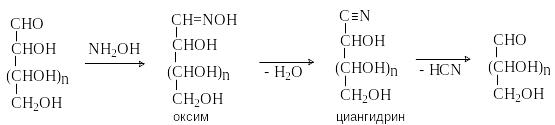 бразование
оксимов и их последующие превращения
позволяют укоротить цепь моносахарида
на один атом углерода. Процесс упрощенно
может быть представлен следующей схемой.
бразование
оксимов и их последующие превращения
позволяют укоротить цепь моносахарида
на один атом углерода. Процесс упрощенно
может быть представлен следующей схемой.
При этом из эпимеров по С-2 образуются одинаковые альдозы (из D-глюкозы и D-маннозы – D-арабиноза)
.
Д еградация по Руффу
Метод состоит в окислении альдозы в гликоновую кислоту с последующим окислительным декарбоксилированием.
Дисахариды
2.1. Строение
Дисахариды состоят из двух моносахаридных остатков, связанных гликозидной связью. Их можно рассматривать как О-гликозиды, в которых агликоном является остаток моносахарида.
Возможно два варианта образования гликозидной связи:
за счет гликозидного гидроксила одного моносахарида и спиртового гидроксила другого моносахарида;
за счет гликозидных гидроксилов обоих моносахаридов.
Дисахарид, образованный первым способом, содержит свободный гликозидный гидроксил, сохраняет способность к цикло-оксо-таутомерии и обладает восстанавливающими свойствами.
В дисахариде, образованном вторым способом, нет свободного гликозидного гидроксила. Такой дисахарид не способен к цикло-оксо-таутомерии и является невосстанавливающим.
В природе в свободном виде встречается незначительное число дисахаридов. Важнейшими из них являются мальтоза, целлобиоза, лактоза и сахароза.
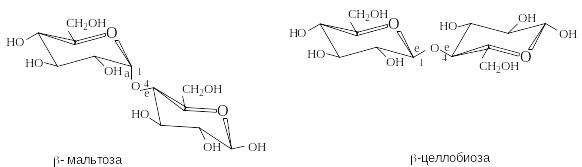
М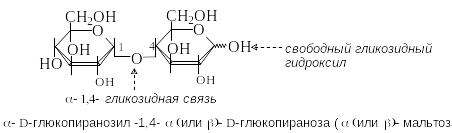 альтоза
содержится в солоде и образуется при
неполном гидролизе крахмала. Молекула
мальтозы состоит из двух остатков
D-глюкозы
в пиранозной форме. Гликозидная связь
между ними образована за счет гликозидного
гидроксила в -конфигурации
одного моносахарида и гидроксильной
группы в положении 4 другого моносахарида.
альтоза
содержится в солоде и образуется при
неполном гидролизе крахмала. Молекула
мальтозы состоит из двух остатков
D-глюкозы
в пиранозной форме. Гликозидная связь
между ними образована за счет гликозидного
гидроксила в -конфигурации
одного моносахарида и гидроксильной
группы в положении 4 другого моносахарида.
М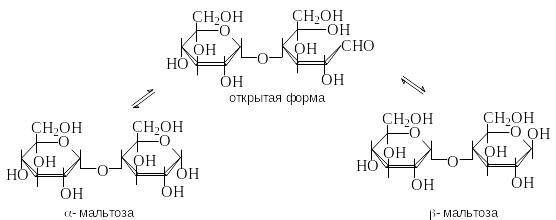 альтоза
– это восстанавливающий
дисахарид. Она способна к таутомерии и
имеет -
и -аномеры.
альтоза
– это восстанавливающий
дисахарид. Она способна к таутомерии и
имеет -
и -аномеры.
Целлобиоза – продукт неполного гидролиза целлюлозы. Молекула целлобиозы состоит из двух остатков D-глюкозы, связанных -1,4-гликозидной связью. Целлобиоза – восстанавливающий дисахарид.
Р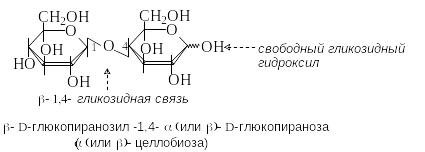 азличие
между мальтозой и целлобиозой состоит
в конфигурации гликозидной связи, что
отражается на их конформационном
строении. Гликозидная связь в мальтозе
имеет аксиальное, в целлобиозе –
экваториальное положение. Конформационное
строение этих дисахаридов служит
первопричиной линейного строения
макромолекул целлюлозы и спиралеобразного
строения амилозы (крахмал), структурными
элементами которых они являются.
азличие
между мальтозой и целлобиозой состоит
в конфигурации гликозидной связи, что
отражается на их конформационном
строении. Гликозидная связь в мальтозе
имеет аксиальное, в целлобиозе –
экваториальное положение. Конформационное
строение этих дисахаридов служит
первопричиной линейного строения
макромолекул целлюлозы и спиралеобразного
строения амилозы (крахмал), структурными
элементами которых они являются.
Л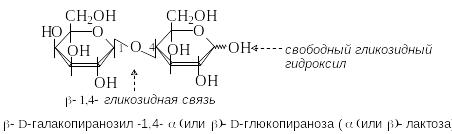 актоза
содержится
в молоке (4-5%). Молекула лактозы состоит
из остатков D-галактозы
и D-глюкозы,
связанных -1,4-гликозидной
связью. Лактоза – восстанавливающий
дисахарид
актоза
содержится
в молоке (4-5%). Молекула лактозы состоит
из остатков D-галактозы
и D-глюкозы,
связанных -1,4-гликозидной
связью. Лактоза – восстанавливающий
дисахарид
С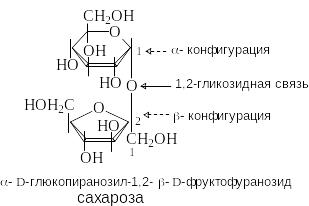 ахароза
содержится в сахарном тростнике, сахарной
свекле, соках растений и плодах. Она
состоит из остатков D-глюкозы
и D-фруктозы,
которые связаны за счет гликозидных
гидроксилов. В составе сахарозы D-глюкоза
находится в пиранозной, а D-фруктоза
– в фуранозной форме. Сахароза –
невосстанавливающий дисахарид.
ахароза
содержится в сахарном тростнике, сахарной
свекле, соках растений и плодах. Она
состоит из остатков D-глюкозы
и D-фруктозы,
которые связаны за счет гликозидных
гидроксилов. В составе сахарозы D-глюкоза
находится в пиранозной, а D-фруктоза
– в фуранозной форме. Сахароза –
невосстанавливающий дисахарид.
2.2. Химические свойства
Дисахариды вступают в большинство реакций, характерных для моносахаридов: образуют простые и сложные эфиры, гликозиды, производные по карбонильной группе. Восстанавливающие дисахариды окисляются до гликобионовых кислот. Гликозидная связь в дисахаридах расщепляется под действием водных растворов кислот и ферментов. В разбавленных растворах щелочей дисахариды устойчивы. Ферменты действуют селективно, расщепляя только - или только -гликозидную связь.
П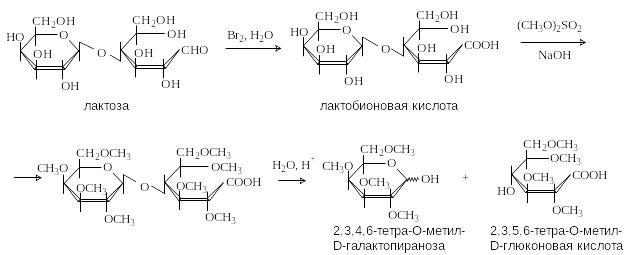 оследовательность
реакций – окисление, метилирование,
гидролиз, позволяет установить строение
дисахарида.
оследовательность
реакций – окисление, метилирование,
гидролиз, позволяет установить строение
дисахарида.
Окисление дает возможность определить, остаток какого моносахарида находится на восстанавливающем конце. Метилирование и гидролиз дают информацию о положении гликозидной связи и размерах цикла моносахаридных звеньев. Конфигурация гликозидной связи ( или ) может быть определена с помощью ферментативного гидролиза.
3. Полисахариды
Полисахариды – полимеры, построенные из моносахаридных остатков, связанных гликозидными связями. Полисахариды могут иметь линейное или разветвленное строение. Полисахариды, состоящие их одинаковых моносахаридных остатков, называют гомополисахаридами, из остатков разных моносахаридов – гетерополисахаридами.
Крахмал – полисахарид растительного происхождения. Его основная биологическая функция – запасное вещество растений. Крахмал представляет собой смесь двух полисахаридов – амилозы (10-20%) и амилопектина (80-90%)
А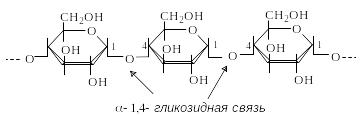 милоза
– линейный гомополисахарид, состоящий
из остатков D-глюкопиранозы,
связанных -1,4-гликозидными
связями. Структурным элементом амилозы
является дисахарид мальтоза.
милоза
– линейный гомополисахарид, состоящий
из остатков D-глюкопиранозы,
связанных -1,4-гликозидными
связями. Структурным элементом амилозы
является дисахарид мальтоза.
Цепь амилозы включает от 200 до 1000 моносахаридных единиц. Вследствие аксиального положения гликозидной связи макромолекула амилозы свернута в спираль.
А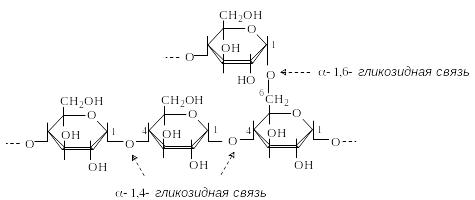 милопектин
– разветвленный гомополисахарид,
построенный из остатков D-глюкопиранозы,
которые связаны в основной цепи
-1,4-гликозидными,
а в местах разветвлений - -1,6-гликозидными
связями. Разветвления расположены через
каждые 20-25 моносахаридных остатков.
милопектин
– разветвленный гомополисахарид,
построенный из остатков D-глюкопиранозы,
которые связаны в основной цепи
-1,4-гликозидными,
а в местах разветвлений - -1,6-гликозидными
связями. Разветвления расположены через
каждые 20-25 моносахаридных остатков.
Крахмал набухает и растворяется в воде, образуя вязкие растворы (гели). Химические свойства крахмала аналогичны свойствам моно- и дисахаридов. Крахмал гидролизуется под действием кислот (но не щелочей) и фермента амилазы. Конечным продуктом гидролиза крахмала является D-глюкоза.
(C6H10O5)n (C6H10O5)m C12H22O11 C6H12O6
крахмал декстрины мальтоза D-глюкоза
n>m
За счет спиралеобразной конформации амилоза способна образовывать соединения включения с молекулярным иодом. Комплексы крахмала с иодом имеют интенсивную синюю окраску. Реакция используется как качественная на иод и крахмал.
Ц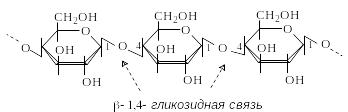 еллюлоза
– самый распространенный растительный
полисахарид. Выполняет функцию опорного
материала растений. Целлюлоза – линейный
гомополисахарид, построенный из остатков
D-глюкопиранозы, связанных
-1,4-гликозидными
связями. Структурным элементом целлюлозы
является целлобиоза.
еллюлоза
– самый распространенный растительный
полисахарид. Выполняет функцию опорного
материала растений. Целлюлоза – линейный
гомополисахарид, построенный из остатков
D-глюкопиранозы, связанных
-1,4-гликозидными
связями. Структурным элементом целлюлозы
является целлобиоза.