Материал: Идентификация генов изоцитратлиазы в прорастающих семенах арабидопсиса
Идентификация генов изоцитратлиазы в прорастающих семенах арабидопсиса
Выпускная работа бакалавра
Биохимия
Идентификация генов изоцитратлиазы в
прорастающих семенах арабидопсиса
Реферат
ГЛИОКСИЛАТНЫЙ ЦИКЛ, ИЗОЦИТРАТЛИАЗА, ГЛЮКОНЕОГЕНЕЗ, ПЦР, ОБРАТНАЯ ТРАНСКРИПЦИЯ, РНК.
Целью работы являлась идентификация генов изоцитратлиазы в прорастающих семенах арабидопсиса. Было установлено, что активность изоцитратлиазы индуцируется в первый день прорастания и постепенно увеличивается до максимумального значения на 4 день. Получены препараты суммарной клеточной популяции РНК. Методом обратной транскрипции получена кДНК. Проведен ОТ-ПЦР анализ, который показал, что две обнаруженные формы фермента являются изоферментами и экспрессируются двумя генами - icl 1 и icl 2.
Список сокращений
кДНК - комплементарная ДНК
ПЦР - полимеразная цепная реакция
РНК - рибонуклеиновая кислота
DTT- дитиотрейтол
EDTA- этилендиаминтетрауксусная кислотаингибитор рибонуклеазы плаценты человека
M-Mlv- ревертаза вируса Moloney лейкоза мышей
Taq- ДНК-полимераза Thermus aquaticus
ИЦЛ - изоцитартлиаза
МС - малатсинтаза
ГЦ - глиоксилатный цикл
ЖК - жирная кислота
ПААГ - полиакриламидный гель
Содержание
Введение
Глава 1. Обзор литературы
.1 Общая характеристика глюконеогенеза
.2 Глюконеогенез в растения
.3 Роль глиоксилатного цикла в глюконеогенез Е
1.4 Характеристика структуры гена и белка ИЦЛ
Глава 2. Экспериментальная часть
.1 Цели и задачи
.2 Объекты и методы исследования
.2.1 Объект исследования
.2.2 Методы исследования
.3 Результаты и их обсуждения
.3.1 Динамика активности изоцитратлиазы
.3.2 Идентификация генов icl 1 и icl 2
Выводы
Литература
Введение
Длительное время подвергается изучению глиоксилатный цикл растений, обеспечивающий растения интермедиатами при конверсии запасных липидов, а также в зеленом растении участвует в превращении токсичных продуктов фотодыхания в безвредные вещества, используемые в конструктивном метаболизме. Ключевым ферментом глиоксилатного цикла является изоцитратлиаза.
Исследование отдельных звеньев клеточного метаболизма является одной из важнейших задач современной биологии. Оно имеет теоретическое и практическое значение, так как позволяет приблизиться к пониманию механизмов функционирования организма.
Изучение
структурно-функциональной организации и молекулярных механизмов регуляции
ключевого фермента глиоксилатного цикла - изоцитратлиазы, является актуальной
проблемой современной биохимии растений.
Глава 1. Обзор литературы
.1 Общая характеристика
глюконеогенеза
Биосинтез глюкозы и других углеводов из более простых предшественников является в количественном отношении наиболее важным процессом в биосфере. Точно также как превращение глюкозы в пировиноградную кислоту, катализируемое ферментами гликолиза, является центральным путем катаболизма углеводов в большинстве клеток как в аэробных, так и в анаэробных условиях, также обратный процесс - превращение пирувата в глюкозу - является наиболее важным общим путем биосинтеза моно- и полисахаридов. В этот центральный биосинтетический путь вливаются два главных «питающих пути»: один из них из ряда реакций, посредством которых интермедиаты ЦТК, лактат, большинство аминокислот, пропионат превращаются в пировиноградную кислоту [рис.1] [этот процесс, протекающий у всех организмов называется глюконеогенезом]; второй важный путь состоит в восстановлении СО2 до глюкозы [этот путь является отличительной особенностью автотрофов].
Глюконеогенез реализуется путем обращения некоторых стадий гликолиза, однако в гликолитическом пути имеются три необратимых стадии, которые не могут использоваться при превращении пирувата в глюкозу, поэтому используются альтернативные реакции [обходные пути], которые термодинамически благоприятствуют синтезу глюкозы.
Первый обходной путь у некоторых животных требует совместного действия цитозольных и митохондриальных ферментов. Реакция, осуществляемая под действием митохондриальной пируваткарбоксилазы, выглядит следующим образом [рис.1]:
глюконеогенез ген белок изоцитратлиаза
ПВК + СО2 + АТФ +
ацетил-КоА = ОАА + АДФ + Фн
Пируваткарбоксилаза -
регуляторный фермент, который в отсутствии ацетил-КоА почти полностью лишен
активности, так как данный метаболит выступает в роли положительного
регулятора. Оксалоацетат, образующийся в этой реакции, восстанавливается, а
затем в митохондриях трансформируется в малат. Малат, диффундируя в цитоплазму
при участии специальной декарбоксилатной транспортной системы, находящейся во
внутренней митохондриальной мембране, окисляется цитоплазматической
НАД-зависимой МДГ с образованием внемитохондриального ОАА.
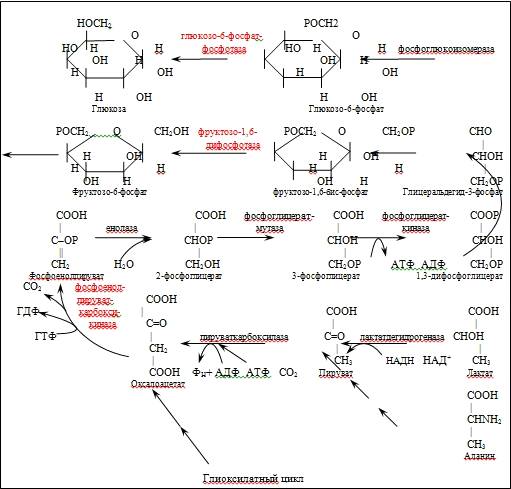
Рис.1. Схема глюконеогенеза: красным выделены ферменты, осуществляющие обходные пути при синтезе глюкозы.
Под действием
фосфоенолпируваткарбоксикиназы [Мn2+
- зависимая реакция, в которой донором фосфата служит ГТФ или АТФ]
щавелевоуксусная кислота распадается на ФЕП и углекислый газ:
ОАА + ГТФ (АТФ) = ФЕП + СО2
+ ГДФ(АДФ)
Фосфофруктокиназная реакция гликолиза необратима, поэтому второй обходной механизм биосинтеза глюкозы достигается при помощи фермента фруктозодифосфотазы, осуществляющим необратимый гидролиз 1-фосфатной группы: Фр-1,6-бис-фосфат + Н2О = Фр-6-Ф + Фн. ∆G0= -3,9ккал/моль.
Для проявления активности фруктозодифосфотазы необходимы ионы Мg2+ и положительный модулятор - АТФ. Данный фермент резко ингибируется отрицательным модулятором - АМФ [5].
На третьем обходном пути вместо гексокиназы, работающей в гликолизе, в глюконеогенезе участвуют глюкозо-6-фосфотаза [Мg2+-зависимый фермент][рис.1].
Таким образом, суммарное
уравнение реакции, ведущей от ПВК к свободной глюкозе следующее:
ПВК + 4АТФ + 2ГТФ + 2НАДН + 6Н2О
= Глюкоза + 2НАД+ +4АДФ +2ГДФ + 6Фн
Субстратами глюконеогенеза выступают также промежуточные продукты ЦТК. Главными из них являются интермедиаты, способные окисляться в малат.
Аминокислоты, служащие предшественниками ФЕП называются гликогенными. Лейцин и некоторые другие аминокислоты являются кетогенными, т.е. способны трансформироваться в ацетоацетат. Фенилаланин, тирозин относятся к смешанным аминокислотам, способным выполнять и кето- и гликогенные функции.
У микроорганизмов и растений
субстратами глюконеогенеза выступают жирные кислоты и ацетил-КоА, способный
вовлекаться в реакции глиоксилатного цикла.
.2 Глюконеогенез в растения
Растение как целостный организм представляет собой сложную саморегулирующуюся систему с взаимозависимыми путями превращения метаболитов. Функционирование и регуляция такой системы обусловлены организацией и биогенезом ферментных комплексов, катализирующих протекание реакций того или иного метаболического пути. Наряду с метаболическими путями, имеющими первостепенную важность в течение всей жизни растений, существуют процессы, протекающие только в определенные периоды онтогенеза. Превращение жиров в углеводы происходит при прорастании семян масличных растений, спор папоротникообразных, при росте пыльцевых трубок и др. [16; 17]. Два первых этапа данного процесса локализованы в глиоксисомах, цикл трикарбоновых кислот - в митохондриях, а последующие два этапа - в растворимой части цитоплазмы.
При прорастании семян запасные жиры, являющиеся водонерастворимыми, превращаются в углеводы, которые хорошо растворяются в воде, что обеспечивает возможность дальнего транспорта запасных веществ к местам утилизиции, где они участвуют в процессах биосинтеза клеточных компонентов. Глиоксилатному циклу принадлежит ключевая роль при глюконеогенезе, поскольку именно наличие ферментов глиоксилатного цикла - изоцитратлиазы и малатсинтазы - обеспечивает протекание этого процесса.
Янтарная кислота, образующаяся в глиоксилатном цикле, используется в дальнейших реакциях глюконеогенеза, а также для регенерации и последующее его окисление до малата и оксалоацетата показаны, в частности, на примере эндосперма клещевины [23]. Последующие две стадии глюконеогенеза, протекающие в цитозоле, изучены пока слабо.
Превращение фосфоенолпирувата в глюкозо-1-фосфат, являющийся ключевым промежуточным соединением в биосинтезе сахаров и полисахаридов, происходит путем обращения части гликолитической последовательности. В связи с тем, что запасные жиры семядолей и других жирозапасающих тканей находятся в особых тельцах - олеосомах, должна иметься тесная связь между олеосомами и глиоксисомами. Эти органоиды не являются производными эндоплазматической сети. Шопфер и др. показали, что имеется тесная связь между мембранами олеосомы и глиоксисомы[22]. В ряде случаев глиоксисома частично «покрывает» олеосому и только тонкий цитоплазматический слой остается между ними. Подобная близость олеосомы и глиоксисомы, по-видимому, обеспечивает стерически-благоприятное состояние для транспорта жирных кислот. Вигил обнаружил, что в клетках эндосперма клещевины глиоксисомы могут располагаться в большом количестве на поверхности более крупных по размеру липидных тел - олеосом[29].
Кислая липаза гидролизует три-,
ди- и моноглицериды, в то время как щелочная липаза расщепляет только
моноглицериды. По-видимому, работа этих двух ферментов взаимосвязана. Однако
данные о локализации липаз, полученные для клещевины, не подтвердились для
других растений. Так, в рапсе [Brassica
napus] липаза не
обнаружена в верхней фракции градиента, представляющей собой олеосомы [19], но
она присутствовала в мембранной фракции, которая, возможно, содержала мембраны
олеосом. Данные, полученные при изучении арахиса [Arachis
hypogaea] [12], указывают
на то, что щелочная липаза связана с мембраной глиоксисом, а также имеется в
митохондриях и пузырьках эндоплазматической сети. Липазная активность не
обнаруживалась во фракции олеосом.
.3 Роль глиоксилатного цикла в
глюконеогенез Е
Глиоксилатный цикл был впервые
открыт Корнбергом и Кребсом в 1957 году в клетках бактерий рода Pseudomonas,
выращивыемых на ацетате [20]. Для этих микроорганизмов было показано, что его
функцией является конденсация С2-молекул для синтеза составляющих
клетку компонентов. Для растительных тканей функционирование глиоксилатного
цикла было впервые показано в эндосперме клещевины [20]. В жирозапасающих
тканях растений основной функцией глиоксилатного цикла также считается участие
в глюконеогенезе при мобилизации запасных жиров. Глиоксилатный цикл шунтирует
две декарбоксилирующие реакции цикла трикарбоновых кислот и позволяет
использовать C2-соединения
в качестве субстрата для глюконеогенеза[3]. Суммарное уравнение глиоксилатного
цикла:
Ацетил-СоА + Н2О +
НАД+ → сукцинат +2СоАSH
+ НАДН + Н+
Глиоксилатный цикл состоит из пяти реакций [рис.2]. На первом этапе ацетил-КоА [C2] конденсируется с глиоксилатом с образованием малата [C4] при помощи ключевого фермента глиоксилатного цикла - малатсинтазы [КФ 4.1.3.2]. НАД-зависимая малатдегидрогеназа окисляет малат до оксалоацетата, восстанавливая НАД+. Цитратсинтаза катализирует образование цитрата из оксалоацетата и ещё одной молекулы ацетил-КоА. Аконитатгидратаза превращает цитрат в изоцитрат через цис-аконитат. Другой маркерный фермент ГЦ изоцитратлиаза [КФ 4.1.3.1] катализирует расщепление изоцитрата на сукцинат и глиоксилат. Другими словами последним этапом глиоксилатного цикла является образование сукцината и регенерация глиоксилата в изоцитратлиазной реакции. Некоторые биохимические этапы ГЦ идентичны реакциям ЦТК, однако существуют их регуляторные особенности, связанные с работой изоферментов и компартментализацией.
Рис.2.Схема реакций
глиоксилатного цикла.
Глюконеогенез, как правило, реализуется при кооперации ферментов, обычно вовлеченных в функционирование различных метаболических путей: липаз, ферментов β-окисления жирных кислот, ферментов глиоксилатного цикла, ЦТК. Липазы обнаружены в олеосомах и глиоксисомах, β-окисление и глиоксилатный цикл протекают в глиоксисомах, окисление сукцината в оксалоацетат [или малат] в митохондриях, конверсия оксалоацетата [малата] в сахарозу - в цитозоле. Реакции ЦТК вносят значительный вклад в глюконеогенез: образовавшийся в ГЦ сукцинат окисляется до малата, затем оксалоацетата, который превращается в фосфоенолпируват с помощью фермента фосфоенолпируваткарбоксикиназы [КФ 4.1.3.32]. Данный фермент, а также фруктозо-1,6-бисфосфатаза [КФ 3.13.11] катализируют два ключевых этапа глюконеогенеза [в отличие от остальных реакций, являющихся простым обращением гликолиза].
Возможность конденсации С2-соединений через глиоксилатный цикл, делает его центральной стадией в пути превращения жирных кислот в углеводы. Основной формой катаболизма жирных кислот [ЖК] является их β-окисление в митохондриях и пероксисомах, приводящее к образованию ацетил-КоА. Как описано выше, две молекулы ацетил-КоА через глиоксилатный цикл могут конденсироваться до сукцината, далее превращающегося в глюкозу.
Процесс превращения ЖК в углеводы имеет место при прорастании семян масличных растений, основным веществом запаспа которых являются липиды. Конверсия запасных липидов в форму углеводов делает возможным построение клеточных компонентов в период, когда интенсивность фотосинтеза низка и необходимы эндогенные источники углеводов [2].
Таким образом, результатом
одного оборота глиоксилатного цикла является синтез одной молекулы глюкогенного
сукцината из двух молекул ацетил-КоА:
ацетил-КоА → сукцинат
Основными биологическими функциями ГЦ является обеспечение роста микроорганизмов на среде с С2-соединениями, синтез клеточных компонентов и конверсия ЖК в углеводы. Кроме того, совсем недавно было открыто, что работа ГЦ является необходимым условием для вируллентности патогенов животных, человека и растений.
У высших растений роль глиоксилатного цикла сводится, как правило к утилизации запасных жиров при прорастании семян и спор. При прорастании семян запасные жиры, являющиеся водонерастворимыми, превращаются в углеводы, которые хорошо растворяются в воде, что обеспечивает возможность дальнего транспорта запасных веществ к местам утилизации, где они участвуют в процессах биосинтеза клеточных компонентов. Глиоксилатному циклу принадлежит ключевая роль при глюконеогенезе, поскольку именно наличие ферментов глиоксилатного цикла - изоцитратлиазы и малатсинтазы - обеспечивает протекание этого процесса.
Пристствие гликсилатного цикла было доказано для прорастающих спор папоротников Dryopteris filixmas, Polypodium vulgare и хвоща Equisetum arvense[14]. У голосеменных также установлено наличие глиоксилатного цикла в прорастающих семенах пихты, некоторых видов сосны[13], а также в гинго. При чем активность глиоксилатного цикла коррелирует со степенью маслянистости семян. ИЦЛ встречается также в созревающих плодах банана[31], в зрелых плодах авокадо[7], в зеленых листьев ряда растений. В настоящее время список покрытосеменных, в которых обнаружен глиоксилатный цикл, достаточно велик.
Долгое время считалось, что ферменты глиоксилатного цикла появляются только при прорастании масличных семян. Но затем было показано, что незначительная активность этих ферментов может наблюдаться и в сухих семенах, при чем показано, что глиоксисомальные ферменты синтезируются уже на стадии созревания семян, присутствуя в микротельцах.
Что касается высших растений, то вопрос о возможности индуцированного синтеза ИЦЛ почти не изучен.
1.4 Характеристика структуры
гена и белка ИЦЛ
Охарактеризован ген ИЦЛ метилотрофных жрожжей Pichia pastoris. Продукт ген ИЦЛ найден скринингом геномной библиотеки P. pastoris. Используя пару вырожденных праймеров, данный ген был клонирован методом ПЦР. Размноженный ген ИЦЛ содержал открытую рамку считывания размером 1563 н. п. Секвенированный белок состоял из 551 аминокислоты[22].
Молекулярная масса субъединицы составила 60,6 кДа с высокой степенью гомологии ИЦЛ из других источников. Аминокислотная гомология составила 64% с ИЦЛ Saccharomyces cerevisiae. Ген ИЦЛ, размером 1458 н.п. был изолирован из метилотрофных дрожжей Hansenula polimorpha. Данная последовательность нуклеотидов кодирует 468 аминокислот белка с молекулярной массой субъединицы 54. 9 кДа.