Материал: Характеристики митохондрий. Гаструляция, ее типы. Понятие лейкоцитов. Биология тимуса
2. Небелковые органические (простетические) группы, принимающие и высвобождающие электроны;
. Белки, обеспечивающие движение электронов.
Каждый из этих комплексов осуществляет перенос электронов от донора к акцептору по градиенту редокс-потенциала через ряд последовательно функционирующих переносчиков. В качестве последних в дыхательной цепи митохондрий функционируют мигрирующие в плоскости мембраны жирорастворимые молекулы убихинона, а также небольшие (молекулярная масса 13 кДа) водорастворимые белки, содержащие ковалентно связанный гем и называемые "цитохромами с". При этом три из пяти компонентов, составляющих дыхательную цепь, работают так, что перенос электронов сопровождается переносом протонов через мембрану крист митохондрий в направлении из матрикса в межмембранное пространство.
Дыхательная цепь начинается с комплекса I (НАДН-убихинон-оксидоредуктаза), состоящего из 16-26 полипептидных цепей и имеющего молекулярную массу около 850 кДа. Функциональная активность этого комплекса определяется тем, что он содержит в своём составе более 20 атомов железа, упакованных в ячейки из атомов серы, а также флавин (Фл - производное витамина рибофлавина). Комплекс I катализирует окисление НАДН, отщепляя от него два электрона, которые после "путешествия" по окислительно-восстановительным компонентам комплекса I попадают на молекулу-переносчик, в качестве которой выступает убихинон (Q). Последний способен ступенчато восстанавливаться, принимая на себя по два электрона и протона и, таким образом, превращаясь в восстановленную форму - убихинол (QH2).
Энергетический потенциал (запас энергии) в молекуле убихинола существенно ниже, чем в молекуле НАДН, а разница в подобной энергии временно запасается в виде особого вида - электрохимического протонного градиента. Последний возникает в результате того, что перенос электронов по простетическим группам комплекса I, ведущий к снижению энергетического потенциала электронов, сопровождается трансмембранным переносом двух протонов из матрикса в межмембранное пространство митохондрии.
Восстановленный убихинол мигрирует в плоскости мембраны, где достигает второго фермента дыхательной цепи - комплекса III (bc1). Последний представляет собой димер из субъединиц b и c1 с молекулярной массой более 300 кДа, сформированный из восьми полипептидных цепей и содержащий атомы железа как в серных ячейках, так и в виде комплексов с гемами b(I), b(II) и c1 - сложными гетероциклическими молекулами с четырьмя атомами азота, расположенными по углам металлосвязывающего квадрата. Комплекс III катализирует реакцию восстановления убихинола до убихинона с передачей электронов на атом железа второй молекулы переносчика (находящегося в межмембранном пространстве цитохрома c). Отщепляющиеся при этом от убихинола два протона водорода освобождаются в межмембранное пространство, продолжая формирование электрохимического градиента. Наконец, ещё два протона водорода переносятся в межмембранное пространство митохондрии за счёт энергии электронов, проходящих по простетических группам комплекса III.
Последняя стадия катализируется комплексом IV (цитохром c-оксидаза) с молекулярной массой около 200 кДа, состоящим из 10-13 полипептидных цепей и, помимо двух различных гемов, включающим также несколько атомов меди, прочно связанных с белками. При этом электроны, отбираемые у восстановленного цитохрома c, пройдя по атомам железа и меди в составе комплекса IV, попадают на связанный в активном центре этого фермента кислород, что приводит к образованию воды.
Таким образом, суммарная реакция, катализируемая ферментами дыхательной цепи, состоит в окислении НАДН кислородом с образованием воды. По сути, этот процесс заключается в ступенчатом переносе электронов между атомами металлов, присутствующих в простетических группах белковых комплексов дыхательной цепи, где каждый последующий комплекс обладает более высоким сродством к электрону, чем предыдущий. При этом сами электроны передаются по цепи до тех пор, пока не соединятся с молекулярным кислородом, обладающим наибольшим сродством к электронам. Освобождаемая же при этом энергия запасается в виде электрохимического (протонного) градиента по обе стороны внутренней мембраны митохондрий. При этом считается, что в процессе транспорта по дыхательной цепи пары электронов перекачивается от трёх до шести протонов.
Завершающим этапом функционирования митохондрии является генерация АТФ, осуществляемая встроенным во внутреннюю мембрану специальным макромолекулярным комплексом с молекулярной массой 500 кДа. Этот комплекс, называемый АТФ-синтетазой, как раз и катализирует синтез АТФ путём конверсии энергии трансмембранного электрохимического градиента протонов водорода в энергию макроэргической связи молекулы АТФ.
Митохондрии и наследственность
Основная статья: Митохондриальные заболевания
ДНК митохондрий наследуются почти исключительно по материнской линии.
Каждая митохондрия имеет несколько участков нуклеотидов в ДНК, идентичных во
всех митохондриях (то есть в клетке много копий митохондриальных ДНК), что
очень важно для митохондрий, неспособных восстанавливать ДНК от повреждений
(наблюдается высокая частота мутаций). Мутации в митохондриальной ДНК являются
причиной целого ряда наследственных заболеваний человека.
. Гаструляция, ее типы
После образования бластулы или морулы в результате перемещения клеточного
материала образуется двухслойный зародыш или гаструла (gaster - желудок).
Процесс, который приводит к образованию гаструлы, называется гаструляцией.
Характерной особенностью гаструляции эмбрионального развития является
интенсивное перемещение клеток, в результате которого будущие зачатки тканей
перемещаются в места, предназначенные для них в соответствии с планом
структурной организации организма. В процессе гаструляции возникают клеточные
слои, которые называются зародышевыми листками. Вначале образуется два
зародышевых листка. Наружный из них получил название эктодермы (ectos - вне,
derma - кожа), а внутренний - энтодермы (entos - внутри). У позвоночных
животных в процессе гаструляции образуется и третий, средний зародышевый листок
- мезодерма (mesos - средний). Мезодерма образуется всегда позже экто- и
энтодермы, поэтому ее называют вторичным зародышевым листком, а экто- и
энтодерму - первичными зародышевыми листками. Эти зародышевые листки вследствие
дальнейшего развития дают начало эмбриональным зачаткам, из которых будут образовываться
различные ткани и органы.
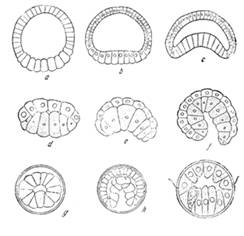
Типы гаструляции
При гаструляции продолжаются изменения, начавшиеся на стадии бластулы, и поэтому разным типам бластул соответствуют и различные типы гаструляции. Переход из бластулы в гаструлу может осуществляться 4-мя основными способами: инвагинацией, иммиграцией, деляминацией и эпиболией.
Инвагинация или впячивание наблюдается в случае целобластулы. Это наиболее простой способ гаструляции, при котором вегетативная часть впячивается в бластоцель. Вначале появляется небольшое углубление в вегетативном полюсе бластулы. Затем клетки вегетативного полюса все больше и больше впячиваются в полость бластоцеля. В последующем эти клетки доходят до внутренней стороны анимального полюса. Первичная полость, бластоцель, при этом вытесняется и видна только с двух сторон гаструлы в местах изгиба клеток. Зародыш принимает куполообразную форму и становится двухслойным. Его стенка состоит из наружного листка - эктодермы и внутреннего - энтодермы. В результате гаструляции образуется новая полость - гастроцель или полость первичной кишки. Она сообщается с внешней средой с помощью кольцеобразного отверстия - бластопора или первичного рта. Края бластопора называются губами. Различают спинную, брюшную и две боковых губы бластопора.
По последующей судьбе бластопора всех животных разделяют на две большие группы: первично- и вторичноротых. К первичноротым относят животных, у которых бластопор остается постоянным или дефинитивным ртом у взрослой особи (черви, моллюски, членистоногие). У других животных (иглокожие, хордовые) бластопор или превращается в заднепроходное отверстие, или зарастает, а ротовое отверстие возникает заново на переднем конце тела зародыша. Таких животных называют вторичноротыми.
Деляминация или расслаивание сводится к расщеплению стенки бластулы. Клетки, которые отделяются внутрь, образуют энтодерму, а наружные - эктодерму. Такой способ гаструляции наблюдается у многих беспозвоночных и высших позвоночных животных.
У некоторых животных в связи с увеличением количества желтка в яйцеклетке и уменьшением полости бластоцеля гаструляция только путем инвагинации становится невозможной. Тогда гаструляция происходит способом эпиболии или обрастания. Этот способ состоит в том, что мелкие анимальные клетки усиленно делятся и обрастают вокруг более крупных вегетативных. Мелкие клетки образуют эктодерму, а клетки вегетативного полюса формируют энтодерму. Такой способ гаструляции наблюдается у круглоротых и земноводных.
Процесс и способы гаструляции
Однако все описанные способы гаструляции редко встречаются отдельно, обычно они комбинируются. Например, вместе с обрастанием может происходить впячивание (земноводные). Расслаивание может наблюдаться вместе с инвагинацией и иммиграцией (рептилии, птицы и др.).
Следовательно, в процессе гаструляции часть клеток из наружного слоя бластулы перемещается внутрь. Это вызвано тем, что в процессе исторического развития одни клетки приспособились к развитию в непосредственной связи с внешней средой, а другие - внутри организма.
На причины гаструляции единого взгляда не существует. Согласно одним взглядам, гаструляция происходит благодаря неравномерному росту клеток в разных частях зародыша. Румблер (1902) объяснял процесс гаструляции изменением формы клеток внутри и снаружи бластулы. Он считал, что клетки имеют клиновидную форму, внутри бластулы шире, а снаружи уже. Существуют взгляды, что гаструляцию может вызывать резкая интенсивность поглощения воды отдельными клетками. Но наблюдения показывают, что эти различия очень невелики.
Гольтфретер (1943) считал, что анимальный полюс бластулы прикрыт тончайшей пленкой (coat) и поэтому клетки связаны в единую массу. Клетки вегетативного полюса не связаны между собой, имеют бутылковидную форму, удлиняются и втягиваются внутрь. В передвижении клеток может играть роль степень слипания и характер межклеточных пространств. Существует также мнение, что клетки могут передвигаться, благодаря их способности к амебоидному движению и фагоцитозу. Образование третьего зародышевого листка в процессе эмбрионального развития животных осуществляется четырьмя способами: телобластическим, энтероцельным, эктодермальным и смешанным.
У многих беспозвоночных животных (первичноротые) мезодерма образуется из двух клеток - телобластов. Эти клетки обособляются рано, еще на стадии дробления. В процессе гаструляции телобласты располагаются на границе между экто- и энтодермой, начинают активно делиться и образующиеся при этом клетки врастают тяжами между наружным и внутренним листками, образуя мезодерму. Такой способ образования мезодермы называется телобластическим.
При энтероцельном способе мезодерма образуется в виде карманообразных
выростов по бокам энтодермы после гаструляции. Эти выпячивания располагаются
между экто- и энтодермой, образуя третий зародышевый листок. Такой способ
образования мезодермы характерен для иглокожих, ланцетника.
3. Функции, строение, классификация лейкоцитов

Лейкоциты (от греч. λευκως - белый и κύτος - клетка, белые кровяные клетки) - неоднородная группа различных по внешнему виду и функциям клеток крови человека или животных, выделенная по признаку отсутствия самостоятельной окраски и наличия ядра.
Главная функция лейкоцитов - защита. Они играют главную роль в специфической и неспецифической защите организма от внешних и внутренних патогенных агентов, а также в реализации типичных патологических процессов. Все виды лейкоцитов способны к активному движению и могут переходить через стенку капилляров и проникать в ткани, где они поглощают и переваривают чужеродные частицы. Этот процесс называется фагоцитоз, а клетки, его осуществляющие, - фагоцитами. Если чужеродных тел проникло в организм очень много, то фагоциты, поглощая их, сильно увеличиваются в размерах и в конце концов разрушаются. При этом освобождаются вещества, вызывающие местную воспалительную реакцию, которая сопровождается отеком, повышением температуры и покраснением пораженного участка. Вещества, вызывающие реакцию воспаления, привлекают новые лейкоциты к месту внедрения чужеродных тел. Уничтожая чужеродные тела и поврежденные клетки, лейкоциты гибнут в больших количествах. Гной, который образуется в тканях при воспалении, - это скопление погибших лейкоцитов.
Количество лейкоцитов
Поскольку число лейкоцитов в крови отражает состояние защитных сил организма, этот показатель интересует врачей всех специальностей. Его определение входит в минимум исследований, которые назначают всем пациентам в стационаре или поликлинике. У здорового человека число лейкоцитов в крови непостоянно. После тяжелой физической работы, приема горячей ванны, у женщин в период беременности, в процессе родов и перед началом менструации оно увеличивается. Это же происходит после приема пищи. Поэтому, чтобы результаты анализа были объективными, его нужно сдавать натощак, утром, не завтракать, можно выпить только стакан воды. В норме содержание лейкоцитов в 1 л крови взрослого человека составляет от 4,0-9,0x109. У детей оно выше: в возрасте одного месяца - 9,2-13,8x109/л, от 1 до 3 лет - 6-17x109/л, в возрасте от 4 до 10 лет - 6,1-11,4x109/л.
Виды лейкоцитов
Лейкоциты различаются по происхождению, функциям и внешнему виду. Некоторые из лейкоцитов способны захватывать и переваривать чужеродные микроорганизмы (фагоцитоз), а другие могут вырабатывать антитела. По морфологическим признакам лейкоциты, окрашенные по Романовскому-Гимзе, со времён Эрлиха традиционно делят на две группы: - зернистые лейкоциты, или гранулоциты - клетки имеющие крупные сегментированные ядра и обнаруживающие специфическую зернистость цитоплазмы; в зависимости от способности воспринимать красители они подразделяются на нейтрофильные, эозинофильные и базофильные. - незернистые лейкоциты, или агранулоциты - клетки, не имеющие специфической зернистости и содержащие простое несегментированное ядро, к ним относятся лимфоциты и моноциты. Ядра зрелых нейтрофильных гранулоцитов имеют перетяжки - сегменты, поэтому их называют сегментоядерными. В незрелых клетках выявляются удлиненные палочковидные ядра - это нейтрофильные палочкоядерные гранулоциты. Еще более "молодые" нейтрофильные гранулоциты носят название "метамиелоциты" ("юные"). Больше всего в крови зрелых сегментоядерных нейтрофильных гранулоцитов, меньше - палочкоядерных, юные формы встречаются редко. По соотношению числа зрелых и незрелых форм можно судить об интенсивности кроветворения. При потере крови для ее восполнения организм начинает продуцировать большое количество клеток. Поскольку они не успевают созреть в костном мозге, в крови появляется много незрелых форм. Сходные процессы происходят при гнойных заболеваниях (аппендицит, перитонит), сепсисе, когда организм старается выработать больше клеток-защитников. При лейкозах лейкоциты начинают размножаться бесконтрольно, поэтому в крови тоже появляется много незрелых форм.
Процентное соотношение отдельных видов лейкоцитов в периферической крови
называется лейкоцитарной формулой. Она рассчитывается на 100 лейкоцитов.
Лейкоцитарная формула позволяет врачу наглядно представить, каких лейкоцитов
много, а каких мало. Изучение лейкоцитарной формулы помогает в определении
степени тяжести инфекционного заболевания, в диагностике лейкозов. Увеличение
числа незрелых нейтрофильных гранулоцитов называется сдвигом лейкоцитарной
формулы влево. Источником лейкоцитов является костный мозг. Облучение,
некоторые лекарственные средства (бутадион, цитостатики, противоэпилептические
препараты) повреждают его. В результате вырабатывается недостаточное число
лейкоцитов, проявляется лейкопения. Увеличение числа лейкоцитов называют
лейкоцитозом, уменьшение - лейкопенией. Наиболее часто лейкоцитоз возникает у
больных с инфекциями (пневмония, скарлатина), гнойными заболеваниями
(аппендицит, перитонит, флегмона), сильными ожогами. Лейкоцитоз развивается в
течение 1-2 ч после начала интенсивного кровотечения. Приступ подагры также
может сопровождаться лейкоцитозом. При некоторых лейкозах число лейкоцитов
возрастает в несколько десятков раз. Хотя проникновение микробов в организм человека
обычно стимулирует иммунную систему, в результате чего число лейкоцитов в крови
увеличивается, при некоторых инфекциях отмечается противоположная картина. Если
защитные силы организма истощены и иммунная система не способна бороться, число
лейкоцитов снижается. Так, например, лейкопения при сепсисе свидетельствует о
тяжелом состоянии больного и неблагоприятном прогнозе. Некоторые инфекции
(брюшной тиф, корь, краснуха, ветряная оспа, малярия, бруцеллез, грипп,
вирусный гепатит) подавляют иммунную систему, поэтому они могут сопровождаться
лейкопенией. Снижение числа лейкоцитов возможно также при системной красной
волчанке, некоторых лейкозах и метастазах опухолей костей.