Материал: Физиология высшей нервной деятельности и сенсорных систем
Механизмы возбуждения кожных рецепторов. Механический стимул приводит к деформации мембраны рецептора. В результате этого электрическое сопротивление мембраны уменьшается, увеличивается ее проницаемость для Na+. Через мембрану рецептора начинает течь ионный ток, приводящий к генерации рецепторного потенциала. При увеличении рецепторного потенциала до критического уровня деполяризации в рецепторе генерируются импульсы, распространяющиеся по волокну в ЦНС.
Рецептивное поле. Совокупность точек на периферии, с которых периферические стимулы влияют на данную сенсорную клетку в ЦНС, называют рецептивным полем.
В одном рецептивном поле находятся рецепторы, посылающие нервные импульсы другим центральным нейронам, т.е. отдельные рецептивные поля перекрываются. Перекрывание рецептивных полей повышает разрешающую способность рецепции и распознавания локализации стимула.
Отношения между интенсивностью стимула и ответом. Существует количественная зависимость между интенсивностью стимула и ответом в форме частоты возникающих потенциалов действия. Такая же зависимость описывает и чувствительность сенсорного нейрона в ЦНС. Различие только в том, что рецептор отвечает на амплитуду стимула, а центральный сенсорный нейрон на частоту потенциалов действия, приходящих к нему от рецептора.
Для центральных сенсорных нейронов важен не столько абсолютный порог S0 стимула, а дифференциальный, т.е. разностный порог. Под дифференциальным порогом понимают минимальное изменение в данном параметре стимула (пространственном, временном и других), которое вызывает измеримое изменение в частоте импульсации сенсорного нейрона. Обычно сильнее всего он зависит от силы стимула. Иначе говоря, чем выше интенсивность стимула, тем выше дифференциальный порог, т.е. тем хуже распознаются различия между стимулами.
Например, для давления на кожу в ограниченном диапазоне некоторых интенсивностей дифференциальный порог равен усилению давления на 3%. Это означает, что два стимула, интенсивности которых по абсолютной величине отличаются на 3% и более, будут распознаваться. Если же их интенсивности отличаются менее чем на 3%, то стимулы будут восприниматься как одинаковые. Следовательно, если после груза в 100 г мы поставим на руку груз в 110 г, то эту разницу мы сможем ощутить. Но если вначале поставить 500 г, а потом - 510 г, то в этом случае разница в 10 грамм не будет распознана, так как она менее 3% (т.е. менее 15 г) величины исходного груза.
Адаптация ощущения. Под адаптацией ощущения понимают уменьшение субъективной чувствительности к стимулу на фоне его непрерывного действия. По скорости адаптации при длящемся действии раздражителя большинство кожных рецепторов разделяют на быстро- и медленно адаптирующиеся. Наиболее быстро адаптируются тактильные рецепторы, расположенные в волосяных фолликулах, а также пластинчатые тельца. Адаптация кожных механорецепторов приводит к тому, что мы перестаем ощущать постоянное давление одежды или привыкаем носить на роговице глаз контактные линзы.
Свойства тактильного восприятия. Ощущение прикосновения и давления на кожу довольно точно локализуется, т. е. относится человеком к определенному участку кожной поверхности. Эта локализация вырабатывается и закрепляется в онтогенезе при участии зрения и проприорецепции. Абсолютная тактильная чувствительность существенно различается в разных частях кожи: от 50 мг до 10 г. Пространственное различение на кожной поверхности, т. е. способность человека раздельно воспринимать прикосновение к двум соседним точкам кожи, также сильно отличается в разных ее участках. На слизистой оболочке языка порог пространственного различия равен 0,5 мм, а на коже спины - более 60 мм. Эти отличия обусловлены главным образом различными размерами кожных рецептивных полей (от 0,5 мм2 до 3 см2) и степенью их перекрытия.
Температурная рецепция [2] . Температура тела человека колеблется в сравнительно узких пределах, поэтому информация о температуре окружающей среды, необходимая для деятельности механизмов терморегуляции, имеет особо важное значение. Терморецепторы располагаются в коже, роговице глаза, в слизистых оболочках, а также в ЦНС (в гипоталамусе). Они делятся на два вида: холодовые и тепловые (их намного меньше и в коже они лежат глубже, чем холодовые). Больше всего терморецепторов в коже лица и шеи. Гистологический тип терморецепторов до конца не выяснен, полагают, что ими могут быть немиелинизированные окончания дендритов афферентных нейронов.
Терморецепторы можно разделить на специфические и неспецифические. Первые возбуждаются лишь температурными воздействиями, вторые отвечают и на механическое раздражение. Рецептивные поля большинства терморецепторов локальны. Терморецепторы реагируют на изменение температуры повышением частоты генерируемых импульсов, устойчиво длящимся все время действия стимула. Повышение частоты импульсации пропорционально изменению температуры, причем постоянная импульсация у тепловых рецепторов наблюдается в диапазоне температуры от 20 до 50 °С, а у холодовых - от 10 до 41 °С. Дифференциальная чувствительность терморецепторов велика: достаточно изменить температуру на 0,2 °С, чтобы вызвать длительные изменения их импульсации.
В некоторых условиях холодовые рецепторы могут быть возбуждены и теплом (выше 45 °С). Этим объясняется возникновение острого ощущения холода при быстром погружении в горячую ванну. Важным фактором, определяющим установившуюся активность терморецепторов, связанных с ними центральных структур и ощущения человека, является абсолютное значение температуры. В то же время начальная интенсивность температурных ощущений зависит от разницы температуры кожи и температуры действующего раздражителя, его площади и места приложения. Так, если руку держали в воде температуры 27 °С, то в первый момент при переносе руки в воду, нагретую до 25 °С, она кажется холодной, однако уже через несколько секунд становится возможной истинная оценка абсолютной температуры воды.
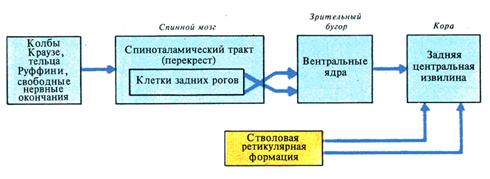
Рис. 4. Блок-схема проводящих путей температурной чувствительности
условный рефлекс кожный чувствительность
В основе периферических нервных механизмов ощущения, в том числе и болевого, лежат сложные взаимодействия различных нервных структур. Ноцицептивный (болевой) импульс, возникающий в рецепторах кожных зон, проводится по аксонам первого нейрона (периферического нейрона), находящегося в клетках межпозвоночных узлов. Аксоны первого нейрона в области задних корешков попадают в спинной мозг и заканчиваются в клетках заднего рога. Следует отметить один важный факт, что на нейронах задних рогов спинного мозга, а также на таламических ядрах (Дуринян Р.А., 1964) конвертируют афферентные волокна кожной чувствительности и болевые афферентные волокна, идущие от внутренних органов. Существенно, при этом, что как соматические, так и вегетативные афферентные волокна оканчиваются не хаотично, а имеют четкую соматотопическую организацию. Эти данные позволяют понять происхождение отраженных болей и зон повышенной кожной чувствительности по Геду при патологии внутренних органов. Второй нейрон, центральный, находится в области заднего рога. Его аксоны, перекрещиваясь в передней спайке, переходят на периферию бокового столба и в составе спинно-таламического пучка достигают зрительного бугра. В области боковых и центральных ядер зрительного бугра, где заканчиваются волокна второго нейрона, находится третий нейрон (тоже центральный), соединяющийся с ядерной зоной коры больших полушарий головного мозга в области задней центральной и теменной извилин. Часть волокон второго нейрона заканчивается в клетках ретикулярной формации ствола мозга, откуда идут волокна третьего нейрона к зрительному бугру.
В процессе фило- и онтогенетического развития кожа из защитного покрова тела стала совершенным органом чувств (Петровский Б.В. и Ефуни С.Н., 1967; Горев В.П., 1967; Есаков А.И. и Дмитриева Т.М., 1971 и др.). Кожный анализатор является особенно удобной моделью для изучения иррадиации, концентрации и индукции нервных процессов (Пшоник А.Т., 1939 и др.). Важное значение в познании механизмов деятельности головного мозга с давних времен имеют пороговые реакции, позволяющие изучать состояние рецепторного аппарата и центральных структур.
Заключение
Физиология высшей нервной деятельности изучает процессы жизнедеятельности организма человека, в основе которых лежит рефлекторная деятельность, позволяющая организму приспосабливаться к меняющимся условиям окружающей среды, адаптироваться к ним и, тем самым выживать - т.е. сохранять свою жизнь и здоровье, под которым понимается не только физическое, но психическое и социальное благополучие.
Физиология высшей нервной деятельности является базовой академической наукой для развития таких практических дисциплин как психология, педагогика, медицина, гигиена труда, спорта, обучения, питания и т.д. Физиология высшей нервной деятельности и свойства нервных процессов определяет и объясняет возрастные и индивидуальные различия поведения человека в постоянно меняющихся условиях внешней среды.
Литература
1. Анатомия и физиология детей и подростков (с возрастными особенностями) / Под ред. Сапина М. Р. - М., 2011
2. Казин Э. М. Основы индивидуального здоровья человека: учебное пособие для вузов - М.: Владос, 2012
. Медведев В. И. Психофизиологические проблемы оптимизации деятельности - М.: Издательский центр «Академия», 2009
. Смирнов В. М. Нейрофизиология и высшая нервная деятельность детей и подростков - М., 2011
. Физиология человека / Под ред. В. М. Покровского - М., 2008