Материал: биохимия . гормоны - 2 часть
Гормоны гипоталамуса
Гипоталамус служит местом непосредственного взаимодействия высших отделов ЦНС и эндокринной системы. Это взаимодействие осуществляется гуморальными факторами – гормональными веществами с чрезвычайно высокой биологической активностью. Эти вещества получили сначала наименование нейрогормонов, а затем рилизинг-факторов (от англ. release – освобождать), или либеринов. Вещества с противоположным действием, т.е. угнетающие освобождение (и, возможно, биосинтез) гипофизарных гормонов, стали называть ингибирующими факторами, или статинами.
В гипоталамусе открыто 7 стимуляторов (либерины) и 3 ингибитора (статины) секреции гормонов гипофиза, а именно:
кортиколиберин,
тиролиберин,
люлиберин,
фоллилиберин,
соматолиберин,
пролактолиберин,
меланолиберин,
соматостатин,
Пролактостатин,
меланостатин
.
В чистом виде выделено 5 гормонов, для которых установлена первичная структура, подтвержденная химическим синтезом.
Гормонов гипоталамуса в исходной ткани содержится чрезвычайно мало. Так, для выделения всего 1 мг тиролиберина потребовалось переработать 7 т гипоталамусов, полученных от 5 млн. овец.
По химическому строению все гормоны гипоталамуса являются низкомолекулярными пептидами, так называемыми олигопептидами необычного строения. Приводим полученные к 1990 году данные о химической природе шести из известных 10 гормонов гипоталамуса.
1. Тиролиберин (Пиро-Глу–Гис–Про–NH2) Тиролиберин представлен трипептидом, состоящим из пироглутаминовой (циклической) кислоты, гистидина и пролинамида, соединенных пептидными связями. В отличие от классических пептидов он не содержит свободных NH2- и СООН-групп у N- и С-концевых аминокислот.
2. Гонадолиберин является декапептидом, состоящим из 10 аминокислот в последовательности: Пиро-Глу–Гис–Трп–Сер–Тир–Гли–Лей–Арг–Про–Гли-NН2
Концевая С-аминокислота представлена глицинамидом.
3. Соматостатин
является циклическим тетрадекапептидом
(состоит из 14 аминокислотных остатков):
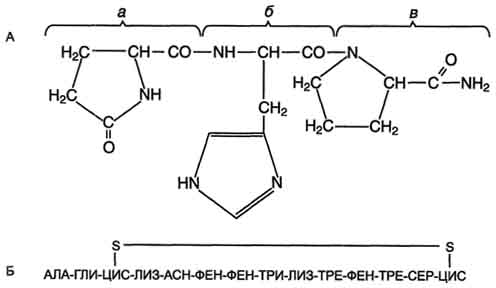
Отличается этот гормон от двух предыдущих, помимо циклической структуры, тем, что не содержит на N-конце пироглутаминовой кислоты: дисульфидная связь образуется между двумя остатками цистеина в 3-м и 14-м положениях. Синтетический линейный аналог соматостатина также наделен аналогичной биологической активностью.
Помимо гипоталамуса, соматостатин продуцируется нейронами центральной и периферической нервных систем, а также синтезируется в S-клетках панкреатических островков (островков Лангерганса) в поджелудочной железе и клетках кишечника.
Он оказывает широкий спектр биологического действия; в частности, ингибирующее действие на синтез гормона роста в аденогипофизе, а также прямое тормозящее действие его на биосинтез инсулина и глюкогона в β- и α-клетках островков Лангерганса.
4. Соматолиберин выделен из природных источников. Он представлен 44 аминокислотными остатками с полностью раскрытой последовательностью. Биологической активностью соматолиберина наделен химически синтезированный декапептид: Н-Вал–Гис–Лей–Сер–Ала–Глу–Глн–Лиз–Глу–Ала-ОН.
Этот декапептид стимулирует синтез и секрецию гормона роста гипофиза соматотропина.
5. Меланолиберин, химическая структура которого (аналогична структуре открытого кольца гормона окситоцина без трипептидной боковой цепи), имеет следующее строение: Н-Цис–Тир–Иле–Глн–Асн–Цис-ОН.
6. Меланостатин (меланотропинингибирующий фактор) представлен или трипептидом: Пиро-Глу–Лей–Гли-NН2, или пентапептидом со следующей последовательностью: Пиро-Глу–Гис–Фен–Aрг–Гли–NН2. Меланолиберин оказывает стимулирующее действие, а меланостатин, напротив, ингибирующее действие на синтез и секрецию меланотропина в передней доле гипофиза.
Помимо перечисленных гипоталамических гормонов, интенсивно изучалась химическая природа другого гормона – кортиколиберина . Активные препараты его были выделены как из ткани гипоталамуса, так и из задней доли гипофиза, которая может служить депо гормона для вазопрессина и окситоцина. Выделен состоящий из 41 аминокислоты .
Местом синтеза гипоталамических гормонов, являются нервные окончания – синаптосомы гипоталамуса, поскольку именно там отмечена наибольшая концентрация гормонов и биогенных аминов.
Гипоталамические гормоны непосредственно влияют на секрецию (точнее, освобождение) «готовых» гормонов и биосинтез этих гормонов de novo. Доказано, что цАМФ участвует в передаче гормонального сигнала. Гормоны гипоталамуса связываются со специфическими аденогипофизарными рецепторами в плазматических мембранах клеток гипофиза, с которыми, после чего через систему аденилатциклазы и мембранных комплексов Са2+–АТФ и Mg2+–АТФ освобождаются ионы Са2+ и цАМФ; последний действует как на освобождение, так и на синтез соответствующего гормона гипофиза путем активирования протеинкиназы.
Гормоны гипофиза
В передней и средней доле гипофиза (аденогипофиз) образуются тропные гормоны. Задняя доля гипофиза (нейрогипофиз) только секретирует гормоны (вазопрессин и окситоцин), которые синтезируются в ядрах гипоталамуса.
В зависимости от места синтеза различают гормоны передней, задней и промежуточной долей гипофиза. В передней доле вырабатываются в основном белковые и полипептидные гормоны, называемые тропными гормонами, или тропинами, вследствие их стимулирующего действия на ряд других эндокринных желез.
Из ткани мозга животных было выделено более 50 пептидов, получивших название нейропептидов и определяющих поведенческие реакции. Эти вещества влияют на некоторые формы поведения, процессы обучения и запоминания, регулируют сон и снимают боль.
Вазопрессин и окситоцин
Гормоны вазопрессин и окситоцин синтезируются рибосомальным путем, причем одновременно в гипоталамусе синтезируются 3 белка: нейрофизин I, II и III, функция которых заключается в нековалентном связывании окситоцина и вазопрессина и транспорте этих гормонов в нейросекреторные гранулы гипоталамуса. Далее в виде комплексов нейрофизин–гормон они мигрируют вдоль аксона и достигают задней доли гипофиза, где откладываются про запас; после диссоциации комплекса свободный гормон секретируется в кровь. Нейрофизины также выделены в чистом виде, и выяснена первичная структура двух из них (92 из 97 аминокислотных остатков соответственно); это богатые цистеином белки, содержащие по семь дисульфидных связей.
Химическое строение обоих гормонов было расшифровано классическими работами американского биохимика Винсента дю Виньо .Оба гормона представляют собой нонапептиды следующего строения:
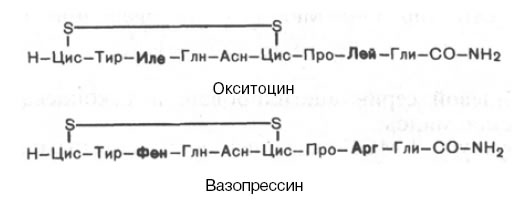
Вазопрессин отличается от окситоцина двумя аминокислотами: он содержит в положении 3 от N-конца фенилаланин вместо изолейцина и в положении 8 – аргенин вместо лейцина. Высказано предположение, что эволюционно все нейрогипофизарные гормоны произошли от одного общего предшественника, а именно аргининвазотоцина, из которого путем одиночных мутаций триплетов генов образовались модифицированные гормоны.
Основной биологический эффект окситоцина у млекопитающих связан со стимуляцией сокращения гладких мышц матки при родах и мышечных волокон вокруг альвеол молочных желез, что вызывает секрецию молока.
Вазопрессин стимулирует сокращение гладких мышечных волокон сосудов, оказывая сильное вазопрессорное действие, однако основная роль его в организме сводится к регуляции водного обмена, откуда его второе название антидиуретического гормона. В небольших концентрациях (0,2 нг на 1 кг массы тела) вазопрессин оказывает мощное антидиуретическое действие – стимулирует обратный ток воды через мембраны почечных канальцев. В норме он контролирует осмотическое давление плазмы крови и водный баланс организма человека. При патологии, в частности атрофии задней доли гипофиза, развивается несахарный диабет – заболевание, характеризующееся выделением чрезвычайно больших количеств жидкости с мочой. При этом нарушен обратный процесс всасывания воды в канальцах почек.
Относительно механизма действия нейрогипофизарных гормонов известно, что гормональные эффекты, в частности вазопрессина, реализуются через аденилатциклазную систему.
Меланоцитстимулирующие гормоны (мсг, меланотропины)
Меланотропины синтезируются и секретируются в кровь промежуточной долей гипофиза. Выделены и расшифрованы первичные структуры двух типов гормонов – α- и β-меланоцитстимулирующие гормоны (α-МСГ и β-МСГ). Оказалось, что у всех обследованных животных α-МСГ состоит из 13 остатков аминокислот, расположенных в одинаковой последовательности: СН3-СО-NH-Сер–Тир–Сер–Мет–Глу–Гис–Фен–Арг–Трп–Гли–Лиз–Про–Вал-СО-NН2
В α-МСГ N-концевой серин ацетилирован, а С-концевая аминокислота представлена валинамидом
Физиологическая роль меланотропинов заключается в стимулировании меланиногенеза у млекопитающих и увеличении количества пигментных клеток (меланоцитов) в кожных покровах земноводных. Возможно также влияние МСГ на окраску меха и секреторную функцию сальных желёз у животных.
Адренокортикотропный гормон (актг, кортикотропин).
Еще в 1926 г. было установлено, что гипофиз оказывает стимулирующее влияние на надпочечники, повышая секрецию гормонов коркового вещества. Накопленные к настоящему времени данные свидетельствуют, что этим свойством наделен АКТГ, вырабатываемый базофильными клетками аденогипофиза. АКТГ, помимо основного действия – стимуляции синтеза и секреции гормонов коры надпочечников, обладает жиромобилизующей и меланоцитстимулирующей активностью.
Молекула АКТГ у всех видов животных содержит 39 аминокислотных остатков. Первичная структура АКТГ свиньи и овцы была расшифрована еще в 1954–1955 гг.
Данные о механизме действия АКТГ на синтез стероидных гормонов свидетельствуют о существенной роли аденилатциклазной системы. АКТГ вступает во взаимодействие со специфическими рецепторами на внешней поверхности клеточной мембраны (рецепторы представлены белками в комплексе с другими молекулами, в частности с сиаловой кислотой). Сигнал затем передается на фермент аденилатциклазу, расположенную на внутренней поверхности клеточной мембраны, которая катализирует распад АТФ и образование цАМФ. Последний активирует протеинкиназу, которая в свою очередь с участием АТФ осуществляет фосфорилирование холинэстеразы, превращающей эфиры холестирина в свободный холестерин, который поступает в митохондрии надпочечников, где содержатся все ферменты, катализирующие превращение холестерина в кортикостероиды.
Соматотропный гормон (стг, гормон роста, соматотропин).
Гормон роста был открыт в экстрактах передней доли гипофиза еще в 1921 г., однако в химически чистом виде получен только в 1956–1957 гг. СТГ синтезируется в ацидофильных клетках передней доли гипофиза; концентрация его в гипофизе составляет 5–15 мг на 1 г ткани, что в 1000 раз превышает концентрацию других гормонов гипофиза. К настоящему времени полностью выяснена первичная структура белковой молекулы СТГ человека, быка и овцы. СТГ человека состоит из 191 аминокислоты и содержит две дисульфидные связи; N- и С-концевые аминокислоты представлены фенилаланином.
СТГ обладает широким спектром биологического действия. Он влияет на все клетки организма, определяя интенсивность обмена углеводов, белков, липидов и минеральных веществ. Он усиливает биосинтез белка, ДНК, РНК и гликогена и в то же время способствует мобилизации жиров из депо и распаду высших жирных кислот и глюкозы в тканях.
СТГ регулирует процессы роста и развития всего организме, что подтверждается клиническими наблюдениями. Так, при гипофизарной карликовости (патология, известная в литературе как пангипопитуитаризм; связана с врожденным недоразвитием гипофиза) отмечается пропорциональное недоразвитие всего тела, в том числе скелета, хотя существенных отклонений в развитии психической деятельности не наблюдается. У взрослого человека также развивается ряд нарушений, связанных с гипо- или гиперфункцией гипофиза.
Известно заболевание акромегалия (от греч. akros – конечность, megas – большой), характеризующееся непропорционально интенсивным ростом отдельных частей тела, например рук, ног, подбородка, надбровных дуг, носа, языка, и разрастанием внутренних органов. Болезнь вызвана, по-видимому, опухолевым поражением передней доли гипофиза.
Лактотропный гормон (пролактин, лютеотропный гормон).
Пролактин считается одним из наиболее «древних» гормонов гипофиза, поскольку его удается обнаружить в гипофизе низших наземных животных, у которых отсутствуют молочные железы, а также получить лактогенный эффект у млекопитающих. Помимо основного действия (стимуляция развития молочных желез и лактации), пролактин имеет важное биологическое значение – стимулирует рост внутренних органов, секрецию желтого тела (отсюда его второе название «лютеотропный гормон»), оказывает ренотропное, эритропоэтическое и гипергликемическое действие и др. Избыток пролактина, образующийся обычно при наличии опухолей из секретирующих пролактин клеток, приводит к прекращению менструаций (аменорея) и увеличению молочных желёз у женщин и к импотенции – у мужчин.
Расшифрована структура пролактина из гипофиза овцы, быка и человека. Это крупный белок, представленный одной полипептидной цепью с тремя дисульфидными связями, состоящий из 199 аминокислотных остатков. Видовые отличия в последовательности аминокислот касаются по существу 2–3 аминокислотных остатков. В гипофизе человека пролактина содержится значительно меньше, чем гормона роста.
В крови женщин уровень пролактина резко повышается перед родами: до 0,2 нг/л против 0,01 нг/л в норме.
Тиреотропный гормон (ттг, тиротропин).
Тиротропин является сложным гликопротеином, выделяемым передней долей гипофиза и содержит по две α- и β-субъединицы, которые в отдельности биологической активностью не обладают.
Полностью расшифрована первичная структура α- и β-субъединиц тиротропина быка, овцы и человека: α-субъединица, содержащая 96 аминокислотных остатков, имеет одинаковую аминокислотную последовательность во всех изученных ТТГ и во всех лютеинизирующих гормонах гипофиза; β-субъединица тиротропина человека, содержащая 112 аминокислотных остатков.
Тиротропин контролирует развитие и функцию щитовидной железы и регулирует биосинтез и секрецию в кровь тиреоидных гормонов. В свою очередь, выделение тиреотропина регулируется по принципу обратной связи гормонами щитовидной железы. Деятельность двух упомянутых желез внутренней секреции тонко координирована..
Гонадотропные гормоны (гонадотррпины).
К гонадотропинам относятся фолликулостимулирующий гормон (ФСГ, фоллитропин) и лютеинизирующий гормон (ЛГ, лютропин), или гормон, стимулирующий интерстициальные клетки. Оба гормона синтезируются в передней доле гипофиза и являются сложными белками – гликопротеинами. Они регулируют стероидо- и гаметогенез в половых желёзах. Фоллитропин вызывает созревание фолликулов в яичниках у самок и сперматогенез – у самцов. Лютропин у самок стимулирует секрецию эстрогенов и прогестерона, как и разрыв фолликулов с образованием желтого тела, а у самцов – секрецию тестостерона и развитие интерстициальной ткани. Биосинтез гонадотропинов регулируется гипоталамическим гормоном гонадолиберином.
Лютропин состоит из двух α- и β-субъединиц. Структура α-субъединиц гормона у большинства животных совпадает. Так, у овцы она содержит 96 аминокислотных остатков и 2 углеводных радикала. У человека α-субъединица гормона укорочена на 7 аминокислотных остатков с N-конца и отличается природой 22 аминокислот. Расшифрована также последовательность аминокислот в β-субъединицах лютропина свиньи и человека. α- и β-Субъединицы в отдельности лишены биологической активности. Только их комплекс, образование которого предопределено первичной структурой их, приводит к формированию биологически активной макромолекулярной структуры за счет гидрофобных взаимодействий.