Материал: био 0411
Ж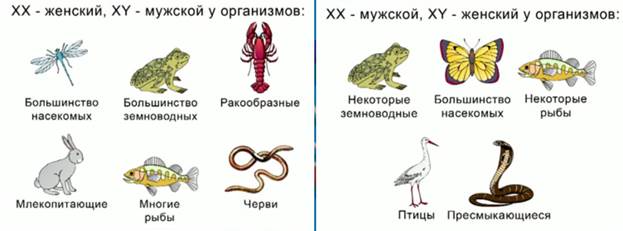 енский
пол производит гаметы Х, мужской пол
производит два типа гамет ХY, в результате
их слияния образуются организмы, в
равной степени несущие женские хромосомы,
ХХ, и организмы, несущие хромосомы
мужские, ХY.
енский
пол производит гаметы Х, мужской пол
производит два типа гамет ХY, в результате
их слияния образуются организмы, в
равной степени несущие женские хромосомы,
ХХ, и организмы, несущие хромосомы
мужские, ХY.
Рождается мальчиков и девочек, в среднем в мире на сто рожденных девочек приходится сто три рожденных мальчика.
Это соотношение с возрастом меняется, к юношеству оно составляет сто на сто, к пятидесяти годам на сто женщин приходится семьдесят пять мужчин, а к восьмидесяти пяти – на сто женщин уже пятьдесят мужчин.
Это вторичное соотношение полов, которое связано с различной жизнеспособностью мужского и женского пола. Человек и млекопитающие животные мужского пола наиболее подвержены влиянию окружающей среды и менее жизнеспособны, поэтому с возрастом соотношение полов меняется.
Для закономерностей Менделя неважно, какой пол несет то или иной признак, они выполняются в любом случае. Но ситуация меняется, если ген находится в половых хромосомах. Y- хромосома генетически инертна, она практически не содержит никаких важных генов для выживания организма, а в Х-хромосоме располагается большое количество генов.
Томас Гент Морган предложил признаки, располагающиеся в Х-хромосоме, назвать признаками, сцепленными с полом.
Рассмотрим влияние гена, находящегося в Х- хромосоме, на его проявление на примере гемофилии (несвертываемости крови).
Здоровая женщина, в роду у которой были гемофилики, вышла замуж за здорового мужчину, какие дети родятся в этом браке?
Н – нормальная свертываемость, h – гемофилия.
X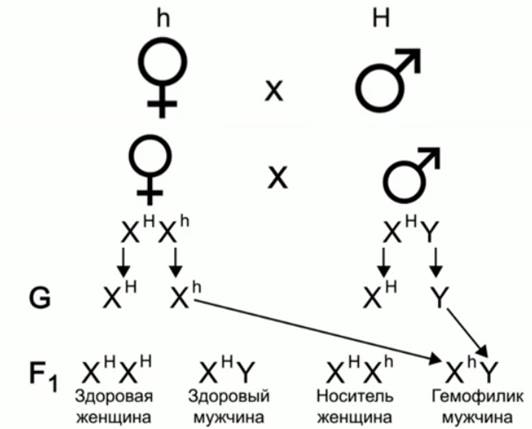 HXh –
генотип матери; ХНY
– генотип отца.
HXh –
генотип матери; ХНY
– генотип отца.
Гаметы, образующиеся в женском организме, – XH и Xh, гаметы, образующиеся в мужском организме, – ХН и Y.
В F1 мы можем видеть следующие генотипы: здоровая женщина, здоровый мужчина, носитель-женщина и гемофилик-мужчина.
Сама женщина здорова, но она гетерозиготна по гену, вызывающему гемофилию, это типичный случай наследования рецессивного признака, сцепленного полом. По такому же принципу наследуется и цветовосприятие человека. Доминантный ген отвечает за нормальное восприятие цвета, а рецессивный ген, который расположен в Х-хромосоме, отвечает за измененное цветовосприятие, называемое дальтонизм, – заболевание, при котором человек не различает красный и зеленый цвета.
Женщины, являясь носителями гена гемофилии и дальтонизма, сами практически не страдают этими заболеваниями, а мужчины страдают этими заболеваниями гораздо чаще.
Вопрос 11 . Что такое сцепление генов? Как наследуются признаки при полном и неполном сцеплении генов? Сцепление генов - это совместное наследование генов, расположенных в одной и той же хромосоме (то есть принадлежат одной группе сцепления и поэтому не могут случайно перекомбинироваться в мейозе, как это бывает при наследовании генов, лежащих в разных хромосомах). Количество групп сцепления соответствует гаплоидному числу хромосом, то есть у дрозофилы 4. Природу сцепленного наследования объяснил. Морган с сотрудниками. В качестве объекта исследования они избрали плодовую муху дрозофилу, которая оказалась очень удобной моделью для изучения данного феномена, так в клетках ее тела, находится только 4 пары хромосом и имеет место высокая скорость плодовитости (в течение года можно исследовать более 20-ти поколений). Сцепленными признаками- признаки, которые контролируются генами, расположенными в одной хромосоме. Они передаются вместе в случаях полного сцепления (закон Моргана). Закон Т. Моргана можно сформулировать следующим образом: гены, находящиеся в одной хромосоме, образуют группу сцепления и часто наследуются совместно, при этом частота совместного наследования зависит от расстояния между генами (чем ближе, тем чаще).
Полное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются так близко друг к другу, что кроссинговер между ними становится невозможным.
Неполное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются на некотором расстоянии друг от друга, что делает возможным кроссинговер между ними.
Полное сцепление встречается редко, обычно – неполное, из-за влияния кроссинговера (перекрещивания и обмена участками гомологичных хромосом в процессе мейоза). То есть, гены одной хромосомы переходят в другую, гомологичную ей.
При полном сцеплении генов А и В по результатам анализирующего скрещивания обнаруживаются два фенотипических класса гибридов, полностью копирующих родителей.
В случае неполного сцепления генов А и В при анализирующем скрещивании появляются четыре фенотипа, два из которых имеют новое сочетание генов: Аb‖аb; аВ‖аb. Появление подобных форм свидетельствует о том, что дигибрид с гаметами АВ│ и аb│ образует кроссоверные гаметы Аb│ и аВ│. Появление таких гамет возможно только в результате обмена участками гомологичных хромосом, то есть в процессе кроссинговера. Количество кроссоверных гамет значительно меньшее, чем некроссоверных.
Сцепленное наследование: два гена находятся в одной хромосоме.
а) При полном сцеплении гетерозигота дает только два типа гамет
б) При неполном сцеплении гетрозигота дает четыре типа гамет, но не с равной вероятностью.
Частота перекреста пропорциональна расстоянию между генами. Чем ближе расположены гены в хромосоме, тем теснее сцепление между ними и тем реже они разделяются при перекресте. И наоборот, чем дальше гены отстоят друг от друга, тем слабее сцепление между ними и чаще перекрест. Следовательно, о расстоянии между генами в хромосомах можно судить по частоте перекреста.
12. Основные положения хромосомной теории Хромосомная теория наследственности — теория, согласно которой передача наследственной информации в ряду поколений связана с передачей хромосом, в которых в определённой и линейной последовательности расположены гены.
Данная теория была сформулирована в начале XX века. Значительный вклад в ее развитие внес американский генетик Томас Морган.
Основные положения хромосомной теории наследственности: 1. Гены расположены в хромосомах. Различные хромосомы содержат неодинаковое число генов. Каждая из не гомологичных хромосом имеет свой уникальный набор генов.
2. Гены расположены в хромосоме линейно.
3. Каждый ген занимает в хромосоме определенный участок (локус). Аллельные гены заполняют одинаковые локусы гомологичных хромосом.
4. Все гены одной хромосомы образуют группу сцепления, благодаря чему происходит сцепленное наследование некоторых признаков. Сила сцепления между двумя генами обратно пропорциональна расстоянию между ними.
5. Сцепления между генами, расположенными в одной хромосоме, нарушаются вследствие кроссинговера, во время которого гомологичные хромосомы обмениваются своими участками.
6. Расстояние между генами в хромосоме пропорционально проценту кроссинговера между ними.
Значение хромосомной теории наследственности для развития биологии заключается в том, что: -С ее помощью выяснено материальная основа законов наследственности, установленная Г. Менделем;
-Закрепила за генами роль элементарных единиц наследственности, локализованных в хромосомах;
-Объясняет отклонения наследования признаков при наличии сцепленного наследования генов.
13.Типы взаимодействия аллельных генов. 1) Полное доминирование-доминантный (преобладающий) ген полностью подавляет действие рецессивного гена при гомозиготном (АА) и гетерозиготном (Аа) наследовании. В фенотипе отражается только один признак, за который отвечает доминантный ген. Однако это не означает, что рецессивный признак полностью исключается. Расщепление признаков по фенотипу во втором поколении будет в соотношении 3:1, т.е. у одной особи полностью проявится рецессивный ген. Свойственно всем живым организмам, включая человека. (Положительный резус-фактор человека всегда доминирует над отрицательным, карий цвет глаз – над голубым, гладкая поверхность горошин доминирует над морщинистым)
2) Неполное доминирование-Промежуточное проявление признаков. Доминантный ген не полностью подавляет действие рецессивного гена. Это отражается на фенотипе в виде неполного проявления признака («усреднение» двух альтернативных признаков). Во втором поколении наблюдается расщепление по генотипу и фенотипу в соотношении 1:2:1 (У ночной красавицы доминантный ген, отвечающий за красный цвет лепестков (АА), не полностью подавляет белый рецессивный ген (аа). В результате получаются гетерозиготные розовые цветки (Аа).
3) Кодоминирование-Полное проявление двух аллельных генов, в фенотипе у гетерозигот одновременно отражаются альтернативные формы признака, доставшиеся от родителей. В этом случае сложно определить доминантный и рецессивный гены, т.к. подавления признака не происходит. (Окраска шортгорнских коров: при скрещивании белых и красных коров получаются пятнистые особи с красно-белой окраской)
4) Множественный аллелизм-Один и тот же ген может иметь в популяции более двух аллелей. Аллельные гены находятся в аналогичных участках хромосом. Множество аллелей обозначается индексами – А1, А2, А3 и т.д. Причина аллелизма – спонтанные изменения гена (мутации), сохраняемые в генофонде в ходе естественного отбора. (Дрозофила имеет 12 вариантов гена, отвечающего за цвет глаз, у человека три аллельных гена определяют группу крови)
5) Аллельное исключение – вид взаимодействия, при котором инактивируется один из аллелей гена, что приводит к проявлению в клетках разных аллелей.
14. Комплементарность- Признак, обусловленный двумя разными генами, проявляется только при сочетании двух доминантных аллелей. Такие гены называются комплементарными. Признак не формируется при отсутствии одного гена. Расщепление фенотипических признаков в F2 происходит в соотношении 9:7, 9:6:1, 9:3:4
Пример.Скрещивание душистого горошка с белыми цветками. В F1 все потомки имеют пурпурные цветки, т.к. сочетание доминантных генов А и В кодируют антоциан, придающий пурпурную окраску. По отдельности гены не образуют пурпур. В F2 происходит расщепление–9 пурпурных (АВ),7белых(3–Abb, 3–aaB, 1–aabb)
Эпистаз. Одна пара генов подавляет другую, не давая проявиться фенотипическому признаку. Подавляющий ген называется эпистатичным (ген-супрессор или ингибитор), подавляемый – гипостатичным. Ингибитор обозначается буквой I, i. Эпистаз может быть доминантным – подавление доминантным геном (I>B, b) и рецессивным – подавление рецессивным геном (i>B,b). При доминировании происходит расщепление генов в соотношении 7:6:3, 12:3:1, 13:3, при рецессивном проявлении – 9:3:4, 9:7, 13
Пример. Окраска овсяного зерна: А – чёрный цвет, В – серый цвет. В F1 все зёрна будут чёрного цвета, если ген А эпистатичен (AaBB или IiBB). В F2 произойдёт расщепление по цвету зерна – 12 чёрных, 3 серых и 1 белое. У 12 растений обязательно присутствует I-ген, у 3 он будет в рецессивном состоянии – i. Одному растению достанутся гены iibb (отсутствие чёрной и серой окраски), поэтому он будет белым
Полимерия. Количественные или мерные признаки, которые нельзя чётко разделить по фенотипу (рост, количество молока, жирность скота), определяются совокупностью генов. Выделяют кумулятивный и некумулятивный виды. В первом случае проявление признака зависит от суммы действий генов (чем больше доминантных генов, тем ярче признак). Во втором случае признак проявляется при доминантном гене, количество генов на проявление фенотипа не влияет. При кумулятивном виде в F2 наблюдается расщепление в соотношении 1:4:6:4:1, при некумулятивном – 15:1. Обозначают полимерные гены одной буквой (А, а, В, b и т.д.), а аллели – цифрой. Например, А1а1А2а2
Цвет кожи человека зависит от действия четырёх генов: А1А1А2А2 – чёрный цвет, а1а1а2а2 – белый, А1А1А2а2, А1а1А2А2, А1а1А2а2, А1А1а2а2, а1а1А2А2, А1а1а2а2, а1а1А2а2 – промежуточные значения от тёмного (почти чёрный) до светлого (почти белый) оттенка
15. Закономерности цитоплазматической наследственности. Цитоплазматическое (или нехромосомное) наследование было открыто в 1909 г. немецкими генетиками Карлом Корренсом (1864–1933) и Эрвином Бауром (1875–1933). Э. Баур первым указал на хлоропласты как на генетические детерминанты изучавшегося им признака пестролистности растений. Генетическая роль митохондрий была открыта в 1949 г. французским генетиком Борисом Эфрусси (1901–1979).
Три закономерности характеризуют цитоплазматическое наследование и отличают его от хромосомного: 1) частое соматическое (митотическое) расщепление;
2) отсутствие или нерегулярное расщепление в мейозе;
3) у многих видов животных и растений имеет место однородительская передача признаков, определяемых структурами цитоплазмы.
Исследования на разных экспериментальных объектах позволили установить три закономерности наследственной передачи, которые отличают цитоплазматические наследственные факторы от хромосомных: 1) соматическое (митотическое) расщепление, именно оно приводит к пестролистности у многих растений;
2) отсутствие расщепления (или нерегулярное расщепление) в мейозе;
3) однородительская передача признака, определяемого цитоплазматическими генами; эта особенность проявляется у самых разных, хотя и не у всех, организмов.
Первые две закономерности обусловлены тем, что в отличие от хромосом цитоплазматические структуры при делении клетки распределяются по дочерним более или менее случайно, во всяком случае нет специального аппарата, который закономерно распределял бы органеллы при клеточном делении.
Третья закономерность – однородительская передача – нередко связывается с тем очевидным обстоятельством, что яйцеклетки гораздо больше по объему своей цитоплазмы, чем спермии, и, соответственно, вносят гораздо больший вклад в образующуюся при слиянии гамет зиготу. Это простое объяснение в большинстве случаев оказывается несостоятельным, его опровергают однородительская передача цитоплазматических генов у изогамной водоросли и отцовская передача хлоропластной или митохондриальной ДНК у некоторых организмов. Механизмы однородительской передачи будут рассмотрены в одном из последующих разделов.
16. Плейотропия (плейотропность) — это множественное действие гена; тип наследования, при котором один ген определяет проявление нескольких признаков.
Виды плейтропии : 1) Первичная: ген одновременно проявляет множественное действие. Например, синдром Марфана обусловлен действием одного гена. Этот синдром проявляется следующими признаками: высокий рост за счёт длинных конечностей, тонкие пальцы (арахнодактилия), подвывих хрусталика, порок сердца, высокий уровень катехоламинов в крови. Другим примером у человека служит серповидноклеточная анемия. Мутация нормального аллеля ведёт к изменению молекулярной структуры белка гемоглобина, при этом эритроциты теряют способность к транспорту кислорода и приобретают серповидную форму вместо округлой. Гомозиготы по гену серповидно клеточности гибнут при рождении, гетерозиготы живут и обладают устойчивостью против малярийного плазмодия. Доминантная мутация, вызывающая у человека укорочение пальцев (брахидактилия), в гомозиготном состоянии приводит к гибели эмбриона на ранних стадиях развития.
2) Вторичная: имеется одно первичное фенотипическое проявление гена, которое обусловливает проявление вторичных признаков. Например, аномальный гемоглобин S в гетерозиготном состоянии фенотипически первично проявляется в виде серповидноклеточной анемии, которая приводит к вторичным фенотипическим проявлениям в виде невосприимчивости к малярии, анемии, гепатолиенальному синдрому, поражению сердца и мозга.
Биологическое значение плейотропии: обеспечивает соотносительный характер наследственной изменчивости, когда изменение одного из признаков сопровождается изменением другого (или нескольких признаков).