Материал: Ассоциативные микроорганизмы тлей
Позже была отмечена тенденция большей выживаемости тли, зараженной комплексом симбионтов Rickettsiella, Hamiltonella, в сравнении с неинфицированными организмами.
Наличие бактерии Regiella
insecticola обеспечивает устойчивость гороховой тли Acyrthosiphon pisium к грибковым патогенам,
таким как Coccidioides immitis и Blastomyces dermatitidis. Это было
экспериментально доказано путем создания изолированной группы тлей,
инфицированных Regiella insecticola, которая впоследствии подверглась заражению
патогенными грибами. По итогам эксперимента был сделан вывод, что данный штамм
в разы увеличивает устойчивость организма-хозяина к патогенным грибам. Так же наличие
симбионта снижает вероятность грибковой споруляции на погибших тлях, как
следствие снижение уровня передачи заболевания другим тлям [21].- это еще один
пример эндосимбионтов. Положительное влияние Wolbachia на организм хозяина
научно доказано. После излечения бактерий рода Wolbachia у организма хозяина
отмечается снижение плодовитости и приспособленности. Бактериями рода Wolbachia
инфицировано, по некоторым оценкам, около 20% всех видов
насекомых.
биологический тля злаковый
1.4.1 Внутриклеточные симбионты тли
К внутриклеточным симбионтам, или эндосимбионтам, относят любой организм, который живет в клетках другого организма, то есть, образует эндосимбиоз. Многие эндосимбионты являются облигатными, т. е. эндосимбионт или хозяин не могут выжить друг без друга. Так же встречаются и факультативные эндосимбионты, присутствие которых в организме хозяина не является строго обязательным.
Эндосимбионты насекомых делятся на две большие группы: «первичные» и «вторичные». Первичные эндосимбионты связаны с организмом хозяев на протяжении долгого периода времени, предположительно от 10 до нескольких сотен миллионов лет. Они образуют облигатные ассоциации. Культивирование таких бактерий, являющихся первичными эндосимбионтами. Жизнеспособными являются в лучшем случае только несколько поколений тлей, живущих без эндосимбионтов.
Вторичные эндосимбионты относятся к не так давно сложившимся ассоциациям. Иногда они горизонтально передаются между организмами хозяев [23].
У некоторых групп насекомых эти эндосимбионты живут в специализированных клетках, называемых бактериоцитами [17]. При созревании половых клеток у самок происходит миграция эндосимбионтов из бактериоцитов в яйцеклетки, другими словами мать передает эндосимбионтов своему потомству. В некоторых случаях, бактерии перемещаются в яйцо, как у Buchnera, у других, таких как Wigglesworthia, они передаются с «молоком» для развивающегося эмбриона насекомых.
Бактерии, в свою очередь, тоже получают выгоду от таких взаимоотношений. Во-первых, на них снижается воздействие хищников. Во-вторых, конкуренция со стороны других видов бактерий отсутствует. В-третьих, бактерии всегда обеспечены достаточным количеством питательных веществ и относительной экологической стабильностью внутри хозяина [21].
Секвенирование генома облигатных эндосимбионтов
насекомых показывает, что их геном гораздо меньше в сравнение с геномом
бактерий, не вступающих в такой тип связи. Бактериальный геном потерял
множество генов в связи с таким существованием. Было выдвинуто несколько
теорий, объясняющих потерю генов. Первое предположение объясняло потерю генов
тем, что существование в щадящих условиях позволило бактериям выключить
некоторые механизмы защиты, и гены, ответственные за работы этих механизмов,
редуцировались. Другая теория предполагает, что относительно небольшое число
бактерий внутри каждого насекомого снизило эффективность естественного отбора,
что привело к потере генов на протяжении многих миллионов лет [24].
1.4.2 Внеклеточные симбионты тли
Помимо внутриклеточных, очень важную роль в жизни хозяев играют внеклеточные симбионты. Исследователи доказали присутствие в пищеварительном тракте тли внеклеточных симбионтов из различных систематических групп эубактерий [21]. Однако в данный момент не до конца выявлены функции вторичных симбионтов и их значение для насекомых. Предполагается их роль в осморегуляции. Тли питаются флоэмным соком, содержащим высокие концентрации углеводов (прежде всего сахарозы и глюкозы), в связи с чем подвергаются постоянному осмотическому стрессу. Известно несколько механизмов, позволяющих насекомым избегать обезвоживания. Высокая концентрация сахарозы непосредственно уменьшает усвоение сахара, синтез олигосахаридов замедляется, тем самым снижается концентрация раствора и, следовательно, осмотическое давление [25 - 28]. Кроме того, в желудок вода дополнительно поступает из кишки, где осмотическое давление ниже.
Недавно был выявлен еще один
механизм осморегуляции тлей - это потребление ксилемного сока растений. Сок
ксилемы является более разбавленным, чем сок флоэмы: концентрация сахаров и
аминокислот в нем не превышает 1%. Дополнительным осморегуляторным механизмом
может служить способность внеклеточных симбионтов тли к использованию сахаров в
качестве источника углерода и энергии, тем самым в процессе своей
жизнедеятельности внеклеточные симбионты понижают концентрацию сахаров в
ксилемном соке, делая его более приемлемым для насекомого [29].
1.5 Тли как вредители
сельскохозяйственных культур
С давних времен известна высокая вредоносность злаковых тлей. Вспышки их массового размножения в России становились причиной гибели посевов или снижения урожаев еще в позапрошлом столетии. И в настоящее время злаковые тли дают вспышки 2-3 раза в 10 лет, потери урожая от них достигают 15 %.
В Западной Сибири условный недобор зерна при сильной степени заселения тлями яровой пшеницы составляет 6 ц/га, урожай с поврежденных посевов содержит до 66 % щуплых семян, что в 6 раз больше, чем с неповрежденных.
На посевах зерновых культур нами отмечено 5 видов злаковых тлей: большая злаковая, черемухово-злаковая, обыкновенная злаковая, волосатая кукурузная. Среди них наиболее массовые виды - большая злаковая и черемухово злаковая тли.
Ущерб, причиняемый вредителями и болезнями растений, велик: мировые потери ежегодно составляют примерно 20 - 25% потенциального мирового урожая продовольственных культур [8 -10].
Процесс циркуляции вирусов тесно связан со способами их передачи, особенностями распространения и сохранения. Знание этих вопросов дает возможность проведения биологически обоснованных мероприятий по защите растений. Важным и малоизученным резервом снижения численности вредных организмов являются регуляторы роста, которые, вызывая физиолого-биохимические изме- нения в растительном организме, способны оказывать определенное влияние на их развитие и без ущерба окружающей среде решить проблему значительного снижения потерь от комплекса вирусов и переносчиков на различных культурах. Физиологически активные вещества существенно влияют на интенсивность процессов образования и оттока пластических веществ из листьев в репродуктивные органы, водный режим, уровень содержания и соотношение компонентов белкового комплекса и углеводов, при этом ингибируя развитие популяций некоторых вредителей. Роль регуляторов роста и развития растений резко возросла в последние годы в связи с широким применением интенсивных технологий возделывания различных культур
Настоящая работа посвящена изучению возможности снижения интенсивности распространения вирусов бобовых культур посредством воздействия фитогормонов на развитие и векторные способности их переносчиков.
Наибольший ущерб урожаю наносят насекомые, что объясняется прежде всего их биологическими особенностями, обилием видов, высокой плодовитостью и быстротой размножения.
Растительноядные насекомые разделяются на многоядных, питающихся растениями разных семейств или питающихся растениями разных видов одного семейства, или одноядных, - преимущественно растениями какого-либо одного вида. Вредных насекомых классифицируют также по группам повреждаемых ими культур - вредители хлебных злаков, вредители овощных культур [15, 16].
Систематическое изменение человеком в процессе сельскохозяйственного производства условий биотопа вызывает соответствующую перестройку биоценологических связей и структуры биоценоза. Образуются так называемые вторичные биотопы и агробиоценозы [15].
С одной стороны, некоторые одноядные виды, питавшиеся ранее на специфичных для целины растениях, погибли, что вызвало обеднение видового состава энтомофауны в новом культурном биотопе, с другой стороны, некоторые виды насекомых, обитавшие раньше на диких злаках, перешли на посевы пшеницы, где нашли изобилие более питательных кормов. Этим в известной степени и объясняется быстрое увеличение численности пшеничного трипса и серой зерновой совки. Предвидеть и регулировать подобные изменения - важнейшая научная и практическая задача [10].
Многие представители подотряда тли являются опаснейшими вредителями растений. Их негативное воздействие заключается в следующем.
Тли повреждают кормовые растения, тем самым истощения, уменьшая объем зеленой массы, плохому плодоношению, преждевременному опадению листьев и образованию галлов, другим видам поражения и деформации частей; в ряде случаев питание тлей заканчивается гибелью растения, происходящей за несколько сезонов.
Повторяя новые и новые витки своего жизненного цикла, тли способны практически бесконечно жить на одном и том же субстрате, вызывая его усыхание. При этом, длительное питание этих насекомых соками растений, содержащими мало белка, не мешает им успешно развиваться и размножаться: у них нет необходимости в белковых веществах, так как, благодаря симбиозу с азотофиксирующими бактериями, они способны усваивать азот прямо из воздуха [8].
В процессе жизнедеятельности тли выделяют сладковатые экскременты - медвяную росу, которую слизывают муравьи. Таким образом, в «лице» муравьев они привлекают на возделываемые участки земли и других вредителей растений. У этих насекомых закреплены механизмы взаимной заботы друг о друге. Тли дают муравьям пищу, а те в ответ оберегают их от хищников, переносят на неповрежденные сочные побеги и даже уносят самок в свои муравейники на зиму. Судя по древним находкам - кусочкам янтаря, в которых находили муравьев вместе с тлями - их взаимовыгодное существование имеет очень длительную историю, поэтому разрушить эту связь и изгнать вредителей бывает сложно, даже используя современные методы защиты растений.
Тли переносят фитопатогенные вирусы,
при этом, один и тот же вид насекомого способен распространять до 100
возбудителей опасных болезней растений [30, 31].
2. Материал и методы
Работа проводилась на базе кафедры
микробиологии и физиологии растений и лаборатории молекулярной биологии
биологического факультета Саратовского государственного университета имени Н.
Г. Чернышевского в период с 2015 по 2016 гг.
2.1 Материал
исследования
Объектом исследования являлась
пшеница сорта Волжанка, Жемчужина поволжья и злаковая тля (Schizaphis graminum)
(рисунок 1), собранная на растениях пшеницы сорта Волжанка и Жемчужина поволжья
в периоды колошения (май), цветения (июнь) и молочной спелости (июль) (рисунок
2,3) на полях ФГБНУ «Научно-исследовательский институт сельского хозяйства
Юго-Востока».
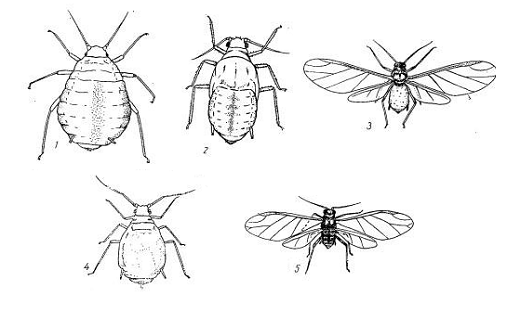
Рисунок 1 - Внешний вид злаковой тля (Schizaphis graminum):
- основательница; 2 - нимфа; 3 - крылатая девственница;
- бескрылая девственница; 5 - самец.

Рисунок 3 - Рожь сорта Волжанка в
периоды колошения (А), налива зерна (Б) и молочной спелости (В).
2.2 Методы исследования
Определение систематического положения насекомых проводили по определителю Блэкман, Истоп [6].
Микробиологические исследования осуществляли стандартными методами. Непосредственно перед бактериологическим посевом насекомых усыпляли, обрабатывали в 70% - ном этаноле в течение пяти мин для уничтожения микроорганизмов, обитающих на внешних покровах, затем промывали в стерильном физиологическом растворе. 10 экземпляров тлей, обработанных таким образом, растирали в ступке с 0,5 мл физиологического раствора. Установлено, что средняя масса 10 особей тлей составляет 0,005 г, таким образом, исходное разведение составляло 10-2. По 0,1 мл полученной суспензии высевали на ГРМ - агар (Россия, Оболенск), картофельную среду (КС) следующего состава: H2O - 1 л, картофель - 200 г, агар - 20 г [36].
Посевы инкубировали при температуре 28ºС в течение 48 - 72 ч. Затем проводили количественный учёт выделенных штаммов микроорганизмов и отсев их на скошенные питательные среды для дальнейшего изучения.
Морфологические и культуральные признаки изучали как на первичных посевах, так и на полученных чистых культурах.
В ходе исследований нами были изучены некоторые биологические свойства микроорганизмов, такие как: способность штаммов к использованию различных сахаров (глюкоза, сахароза, маннит, лактоза, ксилоза, арабиноза, мальтоза, сорбит), цитрата, способность к гидролизу крахмала, желатина, продукции сероводорода, аммиака, индола. При изучении ферментативной активности определяли наличие у выделенных штаммов каталазы и оксидазы.
Так же проводилась работа по определению устойчивости выделенных штаммов к абиотическим факторам. Нами была изучена способность штаммов к росту при различных показателях pH среды (6, 8, 11), концентрациях NaCl в среде (7, 10, 15%), действию температурного фактора (+10 и +43°С).
Определение структуры микробоценозов злаковой тли осуществляли с помощью определения индекса встречаемости и индекса общности различных видов микроорганизмов по В. Н. Беклемишеву [37] с соответствующей модификацией к изучаемым объектам.
Индекс встречаемости рассчитывали как отношение числа проб, в которых обнаружены бактерии данного вида, к общему числу проб, выраженное в процентах.
Идентификацию выделенных штаммов проводили по Определителю бактерий Берджи [38].
Далее штаммы были изучены на наличие ферментов, которые могут найти применение в различных биотехнологических производствах. Нами были выбраны следующие ферменты: пектиназа, целлюлаза и нитрогеназа. Наличие целюлазы определяли путем посева на среду Гетченсона - Клейтона, в состав которой входили (г/л): KH2PO4 - 1,0, MgSO4 - 0,5, FeSO4 - 0,01; NaNO3 - 2,5, карбоксиметилцеллюлоза - 10, агар - 20.
Изучение способности к фиксации молекулярного азота, связанной с наличием фермента нитрогеназы, осуществляли на безазотистой среде Эшби следующего состава (г/л): маннит - 20,0, К2HPO4 - 0,2, MgSO4 - 0,2, NaCl - 0,2, K2SO4 - 0.1, CaCO3 - 5,0, агар - 20.0, pH 7,1 - 7,3.
Постановку всех тестов осуществляли
по общепринятым методикам [39].
3. Результаты исследования
.1 Видовой состав и
численность ассоциативных микроорганизмов злаковой тли
На первом этапе работы нами проводилось изучение основных микробиологических показателей (видовой состав, численность, индекс встречаемости) ассоциативных микроорганизмов злаковой тли, собранных на растении пшеницы: внешняя часть листа, внутренняя часть листа, ризосферы и в самой тле.
В результате проведенных
исследований, из 600 ? особей злаковой тли, собранной в периоды колошения (май)
и молочной спелости (июль) с растений пшеницы и ржи.
Таблица 1. Морфологические и
культуральные признаки микроорганизмов злаковой тли
Виды бактерий
Морфологические признаки
Культуральные признаки
Bacillus
pseudomycoides
1.
2.
Bacillus barbaricus
палочковидные, грамположительные, спорообразующие подвижные
бактерии
Мелкая, непрозрачная, правильная, бесцветная, выпуклая,
блестящая
Bacillus horikoshi
1. Палочка грам + длинные
2. Средняя, правильная, полупрозрачная, цвет(оранж), выпуклая,
блестящая
Bacillus flexus
3. палочковидные, грамположительные, спорообразующие подвижные
бактерии
4. Мелкая, правильная, полупрозрачная, бесцветная, невыпуклая,
блестящая
Bacillus
psychrotolerans
1. палочковидные, грамположительные, спорообразующие подвижные
бактерии
2. Мелкая, правильная, полупрозрачная, бесцветная, невыпуклая,
блестящ
Kurthia sibirica
3. палочковидные, грамположительные, спорообразующие подвижные
бактерии
4. Мелкая, полупрозрачная, правильная, цвет.(ж), выпуклая,
шероховатая
Kurthia gibsonii
5. Бацилла грам +
6. Большая, полупрозрачная, неправильная, бесцветная, плоская,
блестящая
Micrococcus agilis
Кокки, диплококки, тетра, стафилококк
Средняя, непрозрачная, правильная, бесцветная, гладкая, выпуклая
Micrococcus lylae
Кокки грам +
Средняя, полупрозрачная, правильная, бесцветная, выпуклая,
блестящая
Micrococcus
sedentarius
7. Микрококк
8. Мелкая, непрозрачная, правильная, цвет(п), выпуклая, блестящ
Listeria innocua
3. спора
4. Средняя, правильная, непрозрачная, бесцветная, выпуклая,
блестящая
Listeria grayi
Бацилла
Средняя, полупрозрачная, неправильная, цвет(ж), плоская,
блестящая
Таблица 2. Индекс встречаемости и
количественные показатели микроорганизмов в ржи сорта «Волжанка» в период
колошения
Виды
Тля
Внешняя поверхность
листа
Внутренняя среда
Ризосфера
ИВ
КОЕ
ИВ
КОЕ
ИВ
КОЕ
ИВ
КОЕ
Kurthia sibirica
40%
5×103
80%
104
50%
2×103
70%
5×104
Bacillus
pseudomycoides
50%
5×103
70%
5×104
40%
104
20%
5×106
Bacillus barbaricus
50%
104
50%
2×103
20%
5×104
80%
105
Micrococcus agilis
10%
103
60%
3×105
20%
3×103
-
-
Micrococcus lylae
10%
103
100%
105
-
-
-
-
Исследование показало, что в мае в
период колошения, данные микроорганизмы были найдены во всех протестированных
объектах. В тле количественные показатели микроорганизмов варьировали в
диапазоне от 103 до 104 КОЕ в пробе, а максимальная
численность наблюдалась у Kurthia sibirica и Bacillus pseudomycoides. Индекс встречаемости изолированных штаммов находился в пределах
10% - 50%, а максимальный индекс встречаемости выявлен у бактерии Bacillus barbaricus. Количественные
показатели микроорганизмов на внешней части листа варьировали в диапазоне от 2×103 до 5×104 КОЕ в пробе.
Максимальная численность наблюдалась у Bacillus pseudomycoides.
Индекс встречаемости изолированных
штаммов на внешней части листа находился в пределах 50% - 100%. Максимальный
индекс встречаемости был у Micrococcus lylae. Во внутренней части листа количественные показатели
микроорганизмов варьировали от 2×103 до 5×104 КОЕ в пробе. Максимальная численность наблюдалась для Bacillus barbaricus. Индекс встречаемости
изолированных штаммов во внутренней части листа находился в пределах 20% - 50%.
Максимальный индекс встречаемости выявлен у Kurthia sibirica, а так же
микроорганизмы были обнаружены на корнях растения ржи и варьировали от 5×104 до 5×106 КОЕ в пробе.
Максимальная численность наблюдалась у Bacillus pseudomycoides. Индекс встречаемости
изолированных штаммов в ризосфере корнях ржи находился в пределах 20% - 80%.
Максимальный индекс встречаемости выявлен у Bacillus barbaricus.
биологический тля
злаковый
Таблица 3. Индекс встречаемости и
количественные показатели микроорганизмов в ржи сорта «Волжанка» в период
цветения
Виды
Тля
Внешняя пов. листа
Внутренняя среда
Ризосфера
ИВ
КОЕ
ИВ
КОЕ
ИВ
КОЕ
ИВ
КОЕ
9. Kurthia sibirica
50%
104
50%
103
40%
104
80%
5×105
10. Kurthia gibsonii
80%
103
80%
2×103
-
-
50%
4×104
11. Bacillus horikoshi
20%
5×103
20%
2×103
30%
4×105
10%
2×106
12. Bacillus
pseudomycoides
30%
5×103
40%
104
10%
103
20%
106
13. Bacillus
barbaricus
50%
104
20%
3×104
10%
5×103
30%
2×105
14. Bacillus flexus
10%
5×103
80%
2×105
50%
104
10%
105
15. Micrococcus
sedentarius
50%
5×103
20%
2×104
50%
4×104
-
-
16. Micrococcus lylae
40%
103
60%
105
40%
104
20%
103
В период цветения данные
микроорганизмы были найдены во всех протестированных объектах. В тле
количественные показатели микроорганизмов варьировали в диапазоне от 103 до
104 КОЕ в пробе, а максимальная численность наблюдалась у Kurthia gibsonii и Bacillus barbaricus. Индекс встречаемости
изолированных штаммов находился в пределах 10% - 80%, а максимальный индекс
встречаемости был у бактерии Kurthia gibsonii. Количественные показатели микроорганизмов на внешней части листа
варьировали в диапазоне от 103 до 2×105 КОЕ в пробе.
Максимальная численность наблюдалась у Bacillus flexus.
Индекс встречаемости изолированных
штаммов на внешней части листа находился в пределах 20% - 80%. Максимальный
индекс встречаемости были у Kurthia gibsonii и Bacillus flexus. Во внутренней части листа количественные
показатели микроорганизмов варьировали от 103 до 4×105 КОЕ в пробе. Максимальная численность наблюдалась для Bacillus barbaricus. Индекс встречаемости
изолированных штаммов во внутренней части листа находился в пределах 10% - 50%.
Максимальный индекс встречаемости выявлены у Bacillus flexus и Micrococcus sedentarius. Максимальный индекс
встречаемости обнаружены на корнях растения и варьировали от 103 до 2×106 КОЕ в пробе. Максимальная численность наблюдалась у Bacillus
horikoshi. Индекс встречаемости изолированных штаммов в ризосфере корнях ржи
находился в пределах 10% - 80%. Максимальный индекс встречаемости выявлен у Kurthia sibirica.
Таблица 4. Индекс встречаемости и
количественные показатели микроорганизмов в ржи сорта «Волжанка» в период
молочной спелости
Виды
Тля
Внешняя повер. листа Ризосфера
ИВ
КОЕ
ИВ
КОЕ
ИВ
КОЕ
ИВ
КОЕ
5. Kurthia sibirica
20%
103
50%
5×103
40%
5×103
20%
105
6. Listeria innocua
10%
3×103
80%
2×103
80%
5×104
50%
103
7. Listeria grayi
20%
2×103
70%
3×103
50%
103
40%
5×103
8. Bacillus psychrot.
-
-
70%
2×105
-
-
10%
104
9. Bacillus flexus
-
-
60%
103
20%
5×103
20%
103
10. Bacillus horikoshi
-
-
50%
103
20%
5×103
10%
2×104
11. Micrococcus lylae
10%
103
80%
5×103
80%
5×103
50%
104
В период молочной спелости данные
микроорганизмы были найдены во всех протестированных объектах. В тле
количественные показатели микроорганизмов варьировали в диапазоне от 103до
3×103 КОЕ в
пробе, а максимальная численность наблюдалась у Listeria innocua. Индекс встречаемости
изолированных штаммов находился в пределах 10% - 20%, а максимальный индекс
встречаемости были у бактерий Listeria grayi и Kurthia sibirica. Количественные показатели микроорганизмов на внешней части листа
варьировали в диапазоне от 103 до 5×103КОЕ в пробе.
Максимальная численность наблюдалась у Kurthia sibirica и Micrococcus lylae. Индекс встречаемости
изолированных штаммов на внешней части листа находился в пределах 50% - 80%.
Максимальный индекс встречаемости были у Listeria innocua и Micrococcus lylae.
Во внутренней части листа
количественные показатели микроорганизмов варьировали от 103 до 5×104 КОЕ в пробе. Максимальная численность наблюдалась для Listeria innocua. Индекс встречаемости
изолированных штаммов во внутренней части листа находился в пределах 20% - 80%.
Максимальный индекс встречаемости выявлены у Listeria innocua и Micrococcus lylae. Максимальный индекс
встречаемости обнаружены в ризосфере корнях ржи варьировали от 103
до 2×104 КОЕ в пробе. Максимальная численность наблюдалась у Bacillus
horikoshi. Индекс встречаемости изолированных штаммов в ризосфере корнях ржи
находился в пределах 10% - 50%. Максимальный индекс встречаемости выявлены у Listeria innocua и Micrococcus lylae.
Таблица 5. Индекс встречаемости и
количественные показатели микроорганизмов в пшенице сорта «Жемчужина поволжья»
в период колошения
Виды
Тля
Внешняя поверхность листа
Внутренняя среда
Ризосфера
ИВ
КОЕ
ИВ
КОЕ
ИВ
КОЕ
ИВ
КОЕ
Kurthia sibirica
50%
103
80%
2×104
60%
3×103
80%
2×105
Bacillus pseudomycoides
40%
5×103
10%
5×104
10%
104
10%
4×107
Bacillus barbaricus
50%
5×103
70%
5×103
40%
2×104
40%
3×105
Micrococcus agilis
80%
103
40%
105
10%
4×103
-
-
В период колошения данные
микроорганизмы были найдены во всех протестированных объектах. В тле
количественные показатели микроорганизмов варьировали в диапазоне от 103 до
5×103 КОЕ в пробе, а максимальная численность наблюдалась у Bacillus pseudomycoides и Bacillus barbaricus. Индекс встречаемости
изолированных штаммов находился в пределах 40% - 80%, а максимальный индекс
встречаемости был у бактерии Micrococcus agilis. Количественные показатели микроорганизмов на внешней части листа
варьировали в диапазоне от 5×103 до 105 КОЕ в
пробе. Максимальная численность наблюдалась у Micrococcus agilis. Индекс встречаемости
изолированных штаммов на внешней части листа находился в пределах 10% - 80%.
Максимальный индекс встречаемости
был у Kurthia sibirica. Во внутренней части листа количественные показатели
микроорганизмов варьировали от 3×103 до 2×104 КОЕ в пробе. Максимальная численность наблюдалась для Bacillus barbaricus. Индекс встречаемости
изолированных штаммов во внутренней части листа находился в пределах 10% - 60%.
Максимальный индекс встречаемости выявлен у Kurthia sibirica, Максимальный индекс
встречаемости обнаружены на корнях растения и варьировали от 2×105 до 4×107 КОЕ в пробе.
Максимальная численность наблюдалась у Bacillus pseudomycoides. Индекс встречаемости
изолированных штаммов в ризосфере корнях пшеницы находился в пределах 10% -
80%. Максимальный индекс встречаемости выявлен у Kurthia sibirica.
Таблица 6. Индекс встречаемости и
количественные показатели микроорганизмов в пшенице сорта «Жемчужина поволжья»
в период цветения
Виды
Тля
Внешняя поверх. листа
Внутренняя среда
Ризосфера
ИВ
КОЕ
ИВ
КОЕ
ИВ
КОЕ
ИВ
КОЕ
Kurthia gibsonii
80%
103
80%
2×103
50%
103
50%
4×104
Kurthia sibirica
30%
5×103
40%
104
80
104
20%
104
Bacillus horikoshi
20%
2×103
20%
3×103
30%
4×105
10%
2×106
Bacillus pseudomycoides
50%
104
50%
103
40%
104
10%
105
Micrococcus sedentarius
40%
5×103
80%
5×104
50%
2×103
-
-
В период цветения данные
микроорганизмы были найдены во всех протестированных объектах. В тле
количественные показатели микроорганизмов варьировали в диапазоне от 103 до
104 КОЕ в пробе, а максимальная численность наблюдалась у Bacillus pseudomycoides. Индекс
встречаемости изолированных штаммов находился в пределах 20% - 80%, а максимальный
индекс встречаемости был у бактерии Kurthia gibsonii. Количественные показатели микроорганизмов на внешней части листа
варьировали в диапазоне от 103 до 5×104 КОЕ в пробе.
Максимальная численность наблюдалась у Micrococcus sedentarius. Индекс встречаемости
изолированных штаммов на внешней части листа находился в пределах 20% - 80%.
Максимальный индекс встречаемости были у Kurthia gibsonii и Micrococcus sedentarius.
Во внутренней части листа
количественные показатели микроорганизмов варьировали от 103 до 4×105 КОЕ в пробе. Максимальная численность наблюдалась для Bacillus
horikoshi. Индекс встречаемости изолированных штаммов во внутренней части листа
находился в пределах 30% - 50%. Максимальный индекс встречаемости выявлены у Kurthia gibsonii и Micrococcus sedentarius. Максимальный индекс
встречаемости обнаружены на корнях растения и варьировали от 104 до 2×106 КОЕ в пробе. Максимальная численность наблюдалась у Bacillus
horikoshi. Индекс встречаемости изолированных штаммов в ризосфере корнях
пшеницы находился в пределах 10% - 50%. Максимальный индекс встречаемости
выявлен у Kurthia gibsonii.
Таблица 7. Индекс встречаемости и
количественные показатели микроорганизмов в пшенице сорта «Жемчужина поволжья»
в период молочной спелости
Виды
Тля
Внешняя поверхность листа
Внутренняя среда
Ризосфера
ИВ
КОЕ
ИВ
КОЕ
ИВ
КОЕ
ИВ
КОЕ
1. Listeria seeligeri
-
-
50%
5×103
40%
5×103
-
-
2. Listeria grayi
-
-
70%
3×104
20%
103
10%
103
3. Kurthia gibsonii
20%
103
60%
103
20%
5×103
20%
103
4. Bacillus horikoshi
10%
103
80%
5×103
90%
5×104
50%
105
5. Bacillus psychrotolerans
-
-
80%
5×103
10%
105
50%
2×105
6. Micrococcus lylae
20%
2×104
70%
2×105
10%
103
10%
103
В период молочной данные
микроорганизмы были найдены во всех протестированных объектах. В тле
количественные показатели микроорганизмов варьировали в диапазоне от 103 до
2×104 КОЕ в пробе, а максимальная численность наблюдалась у Micrococcus lylae. Индекс встречаемости
изолированных штаммов находился в пределах 10% - 50%, а максимальный индекс
встречаемости был у бактерий Kurthia gibsonii и Micrococcus lylae. Количественные показатели микроорганизмов на внешней части листа
варьировали в диапазоне от 103 до 2×105 КОЕ в пробе.
Максимальная численность наблюдалась у Micrococcus lylae. Индекс встречаемости
изолированных штаммов на внешней части листа находился в пределах 50% - 80%.
Максимальный индекс встречаемости были у Bacillus horikoshi и Bacillus psychrotolerans. Во внутренней
части листа количественные показатели микроорганизмов варьировали от 103
до 105 КОЕ в пробе. Максимальная численность наблюдалась для
Bacillus psychrotolerans. Индекс встречаемости изолированных штаммов во внутренней части
листа находился в пределах 20% - 90%. Максимальный индекс встречаемости выявлен
у Bacillus horikoshi, Максимальный индекс встречаемости обнаружены на корнях
растения и варьировали от 103 до 2×105 КОЕ в пробе.
Максимальная численность наблюдалась у Bacillus psychrotolerans. Индекс
встречаемости изолированных штаммов в ризосфере корнях пшеницы находился в
пределах 10% - 50%. Максимальный индекс встречаемости выявлены у Bacillus
horikoshi и Bacillus psychrotolerans.
Подводя итоги по выше изложенному
можно сказать, что многие виды микроорганизмов, такие как Bacillus horikoshi,
Bacillus psychrotolerans, Kurthia sibirica хорошо распространены во всех отделах растения. Это говорит о
том, что данные микроорганизмы имеют благоприятные условия для их
существования.
3.2 Биологические
свойства бактерий, выделенных из злаковой тли
В ходе постановки биохимических
тестов нами были проведены исследования сахаролитических свойств
микроорганизмов - ассоциантов злаковой тли (таблица 3). Результаты показали,
что все исследуемые виды способны расщеплять только 16,6% глюкозы и 25%
ксилозы, 41,6% видов утилизируют маннит и 33,6% сахарозу. Полученные данные
согласуются с данными о составе флоэмного сока, которым могут питаться злаковые
тли. В его состав входят различные углеводы [42].
Способность к редукции нитратов
проявляли все изолированные микроорганизмы - ассоцианты, что также связано с
особенностями питания насекомых. Основной источник питания злаковой тли -
ксилемный сок, в состав которого входят нитраты. Источником органического азота
для исследуемых насекомых является флоэмный сок, однако содержание аминокислот
в нем достаточно низкое [41]. Недостаток органического азота тли восполняют за
счёт азотофиксирующих ассоциативных микроорганизмов, к процессу фиксации
молекулярного азота были способны все изолированные виды бацилл.
Так же нами был изучен диапазон
устойчивости выделенных из тли бактерий к некоторым физико-химическим факторам:
температура, pH, концентрация NaCl в среде (таблица 4). Выявлено что, щелочное значение pH при 7% предпочитают 50%
изолятов, а при pH 10%, только 25%. Культивирование в 10°С было благоприятно для
66,6% изолятов, а для при 43°С было выделено в 50% исследуемых штаммов. Наличие
15% NaCl в среде стало летальным для 100% исследуемых штаммов.
Таблица 8. Биохимическая активность
микроорганизмов злаковой тли
Таблица 9 Устойчивость
микроорганизмов злаковой тли к физико-химическим факторам
Название теста
Положительный результат, %
Отрицательный результат, %
Температура культивирования
10°С
66,6
33,3
43°С
50
50
Концентрация NaCl
7%
50
50
10%
25
75
15%
0
0
Диапазон pH
5
0
0
6
75
25
9
83,3
16,7
Диапазон pH
10
58,3
41,7
Таблица 10. Целлюлолитическая
активность выделенных штаммов
Виды бактерий
Целлюлолитическая активность
Bacillus pseudomycoides
+
Bacillus barbaricus
+
Bacillus horikoshi
+
Bacillus flexus
+
Bacillus psychrotolerans
+
Kurthia sibirica
-
Kurthia gibsonii
-
Micrococcus agilis
+
Micrococcus lylae
+
Micrococcus sedentarius
-
Listeria innocua
+
Listeria grayi
+
биологический тля
злаковый
Изучение способности к фиксации
молекулярного азота осуществлялась на безазотистой среде Эшби. (таблица 7). К
данному процессу были способны не все исследуемых штаммов (таблица 7), а именно
виды Kurthia и Micrococcus sedentarius
Таблица 11. Способность к фиксации
молекулярного азота
Виды бактерий
Фиксация N2
Bacillus pseudomycoides
+
Bacillus barbaricus
+
Bacillus horikoshi
+
Bacillus flexus
+
Bacillus psychrotolerans
+
Kurthia sibirica
+
Kurthia gibsonii
-
Micrococcus agilis
+
Micrococcus lylae
-
Micrococcus sedentarius
- +
Listeria grayi
+
Таким образом, наши исследования
показали, что организм злаковой тли является средой обитания для узкого круга
сапрофитических бактерий. Насекомое обеспечивает этих бактерий пищевым
субстратом, сглаживает негативное влияние факторов внешней среды. Ассоциативные
бактерии же, в свою очередь, могут участвовать в решении ряда физиологических
проблем организма хозяина, связанных с питанием растительными соками.
Заключение
Микробиоценоз тлей - это активная
динамическая структура. Бактерии - ассоцианты могут снижать осмотическое
давление в пищеварительном канале тли за счёт активного потребления сахаров,
содержащихся в растительном соке. Так же ассоциативные бактерии могут обогащать
организм насекомого-хозяина азотом за счет фиксации молекулярного азота.
Изучение микробоценозов тли с
растениями - это важный этап разработки новых микробиологических методов
ограничения численности насекомых. Существующих биопрепаратов на основе
облигатных энтомопатогенов крайне мало и они имеют ряд недостатков. Причина
этого в высокой степени специализации таких микроорганизмов: гибель насекомых
влечет за собой отмирание самих возбудителей бактериозов.
Список использованных источников
1. Douglas, A. E. Symbiotic interaction / A. E.
Douglas. Oxford: Univer. Press, 1994. 148 p.
2. Маргелис, Л. Роль симбиоза в эволюции клетки / Л.
Маргелис. М.: Мир, 1983. 354 с.
3. URL: http://www.biology-online.org/dictionary/Symbiosis. Дата обращения: 18.03.2015
. Бухарин, О. В. Ассоциативный симбиоз / О. В. Бухарин, . Проворов, Н. А. Генетические и молекулярные основы
симбиотических адаптаций / Н. А. Проворов, И. А. Тихонович // Успехи 6. Blackman, R. L. Aphids on the World's Herbaceous Plants
and Shrubs / R. L. Blackman, V. F. Eastop. St. Louis: Daya Publishing House,
2006. 1439 p.
. Favret, C. Catalog of the aphid genera described from the New World / C. Favret, G. L.
Miller, Nieto Nafría, F. Trans // Soc. Am. 2008. Vol. 133. P. 363 - 412.
8. Васильев, В. П. Вредители плодовых культур / В. П.
Васильев. М.: Государственное издательство сельскохозяйственной литературы, 1958.
399 с.
. Бровдий, В. М. Вредители сельскохозяйственных культур /
В. М. Бровдий. Киев: Урожай, 1973. 576 с.
. Горбачев, И. В. Защита растений от вредителей / В. В.
Горбачев, И. В. Гриценко, Ю. А. Захваткин. М.: Колос, 2002. 472 с.
. Мордвилко, А. К. Насекомые полужесткокрылые (Insecta
Hemiptera). Aphidodea. Фауна России и сопредельных стран, преимущественно по
коллекциям Зоологического Музея Императорской Академии Наук / А. К. Мордвилко.
Петроград: Типография Российской Академии Наук, 1919. Т. 1. 276 с.
12. Comstock, J. H. The Wings of Insects / J. H. Comstock.
New-York: Comstock Publishing, 1918. 430 p.
. Heie, O. E. Palaeontology and phylogeny. Aphids: Their
biology, natural enemies, and control / O. E Heie, A. K. Minks, P. Harrewijn.
Amsterdam: Elsevier, 1987. 568 p.
. Minks, A. K. Aphids: Their biology, natural enemies and
control / A. K. Minks, P. Harrewijn. Amsterdam: Elsevie, 1987. 485 p.
15. Дамрозе, И. П. Эколого-таксономическая и темпоральная
обусловленность распределения популяций крылатых тлей / И. П. Дамрозе. Елгава:
Москва, 1990. 180 с.
. Бобырь, А. Д. Защита растений от вредителей и болезней и
терапия вирусных болезней / А. Д. Бобырь. Киев: Наукова думка, 1962. 566 с.
17. Douglas, A. E. Nutritional Interactions in Insect-Microbial
Symbioses: Aphids and Their Symbiotic Bacteria Buchnera / A. E. Douglas //
Annual Review of Entomology. 1998. Vol. 43. P.
17 - 37.
18. Sauer, C. Bakterielle Endosymbiosen in Insekten / C. Sauer,
19. Hampton, O. R. The nature of bean yield reduction by bean
yellow andmosak viruses / O. R. Hampton // Annals of the Entomological Society
of America. 1975. Vol. 65. P. 1342 - 1346.
20. Kennedy, J. S. Benefits to aphids from feeding on galled
and virus-infected leaves / J. S. Kennedy // Nature. 1953. Vol. 195. P. 168 -
825.
21. Baumann, P. The prokaryotes / P. Baumann, N. A. Moranl. New-York: Springer, 2000. 526 p.
22. Tsuchida, Y. R. Distribution and diversity of Wolbachia in
different populations of the wheat aphid Sitobion miscanthi / Y. R. Tsuchida // Global Perspectives on Biology and Management. 2010.
Vol. 37. P. 265 - 272.
23. Van Der Wilk, F. Isolation and characterization of APSE -
1, a bacteriophage infecting the secondary endosymbiont of Acyrthosiphon pisum
/ 24. Dunbar, H. E. Aphid Thermal Tolerance Is Governed by a
Point Mutation in Bacterial Symbionts / H. E. Dunbar, A. C. Wilson, N. R.
Ferguson, . Wilkinson, T. L. Honeydew sugars and osmoregulation in
the Pea aphid Acyrthosiphon pisum / T. L. Wilkinson, D. A. Ashford, J.
Pritchard, A. E. Douglas // Journal of Experimental Biology. 1997. Vol. 11. P. 2137 - 2143.
26. Ashford, D. A. Living on a high Sugar diet: the fate of
sucrose ingested by a phloem feeding insect, the Pea aphid Acyrthosiphon pisum
/ D. A. Ashford, W. A. Smith, A. E. Douglas // Journal of Insect Physiology. 2000. 27. Malone, M. The spittlebug Philaenus spumarius feeds from
mature xylem at the full hydraulic tension of the transpiration stream / M.
Malone, R. Watson, J. Pritchard // New Phytologist. 1999. Vol. 143. P. 261 - 271.
28. Powell, G. Xylem ingestion by winged aphids / G. Powell, J.
Hardie // Entomologia Experimentalis et Applicata. 2002. Vol. 104. P. 103 - 108.
29. Pompon, J. Role of xylem consumption on osmoregulation in
Macrosiphum euphorbiae / J. Pompon, D. Quiring, P. Giordanengo, Y. Pelletier //
Journal of Insect Physiology. 2010. Vol. 56. P.
610 - 615.
Е. С. Лобакова, Н. В. Немцева, С. В. Черкасов. Екатеринбург: УрО РАН, 2007. 264
с.
современной биологии. 2014. Т. 134, № 3. С. 211 - 226.
B. Hölldobler, R. Gross // Biospektrum. 2006.
Vol. 5. P. 359 - 363.
F. Van Der Wilk, A. M. Dullemans, M. Verbeek, J. F. van den Heuvel // Virology.
1999. Vol. 15, №1. P. 104 - 108.
N. A. Moran // PLoS Biol. 2007. Vol. 15. P. 231 - 242.
Vol. 46. P. 335 - 343.