Материал: Ассоциативные микроорганизмы тлей
Большую часть бактериального населения семян всех растений составляют неспороносные палочки из рода Pseudomonas, активно размножающиеся на поверхности растений. Особенно часто встречается Ps. herbicola, которая образует на твердых питательных средах колонии золотисто-желтого цвета. Преобладание этой бактерии па зерне служит показателем его хороших качеств. При всевозможных пороках этот микроорганизм вытесняется другими. Нередко на зерне может быть найден Ps. fluorescens, дающий флюоресцирующие колонии. Из бактерий других видов на зерно встречаются микрококки, молочнокислые бактерии и т. д. На зернах кукурузы после удаления оберток несколько увеличивается число микрококков. По этому признаку микрофлора кукурузы несколько отличается от микронаселения других зерновых культур.
На зерне встречаются и другие разнообразные представители бактериального мира, попадающие с пылью и переносимые насекомыми. На поверхности зерна они сохраняются в пассивном состоянии и поэтому их число невелико. К отмеченной категории можно отнести маслянокислые бактерии и большинство бацилл.Грибов на поверхности растений и на зерне немного, вероятно, их размножению препятствует слабая концентрация питательных веществ.
Обычно грамм доброкачественного зерна содержит несколько тысяч зародышей грибов.
Итак, можно сказать, что эпифитная грибная микрофлора зерна не столь многочисленна. В ней обычно встречаются виды рода Penicillium, составляющие нередко большинство грибных организмов. Род Aspergillus представлен на здоровом зерне менее обильно. В зерне южного происхождения роль грибов рода Aspergillus возрастает, так как почвы юга более богаты грибами этого рода. В значительно меньшем количестве на зерне встречаются другие грибы, принадлежащие к родам Gladosporium, Alternaria, Mucor и т. д. Здесь же могут быть найдены и дрожжи.
В отличие от других злаков, на свежеубранном зерне кукурузы встречаются грибы из родов Cephalosporium и Fusarium. При хранении зерна количество этих грибов сильно уменьшается.
Актиномицеты не являются существенным компонентом микрофлоры зерна и видовой состав их на этом субстрате пока по привлекал внимания исследователей.
Эпифитная микрофлора зерна находится на поверхности зерна и в нормальной обстановке не проникает в его ткани. Болезнетворные микроорганизмы могут внедриться в зерно, которое становится впоследствии источником распространения инфекции.
Фитопатологи обычно обращают внимание только на резко проявляющиеся болезни растений. Между тем имеются скрытые поражения, приносящие тем не менее значительный урон сельскому хозяйству.
Грибы из рода Alternaria, переходя с корневой системы растений на надземную часть, проникают в ткани растения, сильно ослабляют ого рост и снижают урожай. Подобные грибы по существу вызывают скрытые заболевания растений. Это говорит о необходимости бороться не только с явными, но и скрытыми болезнями растений.
По своему химическому составу зерно является весьма полноценной питательной средой для микроорганизмов. Трудность проникновения внутрь зерна метатрофных микробов объясняется тем, что у него имеются защитные средства. Здесь действует механический и активный иммунитет растения.
У созревающего зерна довольно большая влажность и на его поверхность путем экзосмоса поступают некоторые питательные вещества. Это дает возможность микроорганизмам развиваться на поверхности зерна. Энергичному экзосмосу содействуют и интенсивные биохимические процессы, протекающие в зерне на ранних стадиях его развития.
К моменту полного созревания в зерне уменьшается содержание более активных форм углеводов, но возрастает запас крахмала и гемицеллюлез.
Падает активность гидролаз и дыхания, снижается энергия и других ферментативных процессов. Особенно важно то, что процесс созревания зерна связан с большой потерей воды. Зерно в стадии молочной спелости содержит около 50% влаги; в момент восковой спелости влажность падает до 25 %, при полной до 12%. Микроорганизмы могут размножаться лишь на субстратах, достаточно богатых водой. Вполне созревшее зерно не может считаться таким субстратом, так как вода в нем находится в связанной форме. На спелом сухом зерне микробы сохраняются в покоящемся состоянии (анабиозе), при этом их численность постепенно снижается. В случае подмочки зерна или его увлажнения от других причин микроорганизмы в зерне начинают бурно размножаться и вызывают его порчу.
Отдельные группы микрофлоры начинают развиваться на зерне при разных уровнях влажности. Так, при температуре около 15- 20°С некоторые грибы могут размножаться на зерне пшеницы и кукурузы с влажностью 14,5-15%, а бактерии - при увлажнении зерна пшеницы до 17,5-18%. Для зерна различных культур имеется своя критическая влажность, при которой на нем возможно размножение микроорганизмов. На семенах бобов при указанной выше температуре грибы развиваются при влажности 16%, подсолнечника- 7-9%. Это зависит от количества связанной воды у различных семян, что определяется их структурой и химическим составом. Микроорганизмы начинают развиваться на зерне, лишь, когда в нем появляется свободная вода, то есть степень увлажнения превышает уровень связанной воды.
Степень увлажнения хранящегося зерна зависит от влажности окружающего воздуха, К настоящему времени установлены значения равновесной влажности семян и зерна растений при различной влажности воздуха. Руководствуясь этими показателями, можно создавать благоприятные условия для хранения зерна и семян.
Развитие микроорганизмов на зерне и семенах зависит также от температуры. Эта зависимость отмечается только для несколько увлажненного материала, так как на сухом зерне независимо от температуры микроорганизмы не развиваются. При повышенной влажности зерна микроорганизмы размножаются тем быстрее, чем выше температура.
При температуре 100С даже довольно влажное зерно (18-19% влаги) может хорошо храниться, а при 15-20°С оно начинает быстро плесневеть и портиться бактериями. Для успешного хранения зерна при более высокой температуре его влажность необходимо снизить.
Активное развитие микроорганизмов в зерновой массе различных культур при одной и той же степени увлажнения начинается в разные сроки. Пшеница, рожь, ячмень, горох, бобы и гречиха более устойчивы. В просе, кукурузе и подсолнечнике микроорганизмы развиваются быстрее и интенсивнее.
Следовательно, по составу микрофлоры зерна можно судить об условиях его хранения.
При более или менее длительном развитии микроорганизмов в результате их жизнедеятельности масса зерна может разогреться. Зерновые массы имеют низкую теплопроводность и поэтому хорошо аккумулируют тепло. Наряду с микроорганизмами тепло выделяется вследствие дыхания зерна, развития насекомых и т. п. Глубоко зашедший процесс разогревания зерна приводит к повышению его температуры до 60°С. Зерно при этом нередко приобретает темную окраску - «обугливается», так как в нем образуются темноокрашенные соединения меланоидной природы.
Сохранность урожая
овощей и плодов, имеющих большую влажность, определяется их иммунитетом и
созданием внешней среды, предупреждающей развитие микроорганизмов на их
поверхности.
1.4 Эндофитная
микрофлора растений
В последнее десятилетие большинство сельскохозяйственных ориентированных исследований было направлено на изучение ризосферных микроорганизмов. Среди них бактерии являются наиболее технологичными как с точки зрения производства, так и применения микробных препаратов в сельском хозяйстве. Это открывает перспективы использования прокариоти-ческих микроорганизмов, благотворно влияющих на развитие растений в различных условиях.
Эндофитные бактерии колонизируют те же экологические ниши в растении, что и болезнетворные микроорганизмы, и поэтому являются перспективным биологическим средством для борьбы с фитопатогенами, то есть являются так называемым "биоконтрольным" агентом. Установлено, что бактериальные эндофиты способны ингибировать развитие насекомых-фитофагов и нематод, подавляя их действие путем синтеза биологически активных соединений, обладающих "антипатогенным" действием.
Изучение "биохимического оружия" таких бактерий позволит выделить и идентифицировать те химические соединения, которые могут быть использованы при создании новых препаратов для борьбы с болезнями растений, животных и даже человека.
Некоторые штаммы эндофитных бактерий нашли применение при фиторемедиа ции почв, то есть очистке техногенно загрязненных территорий путем создания специальных растительно-бактериальных систем].
В обзоре систематизированы литературные данные об эндофитных бактериях и их использовании при разработке новых микробных препаратов, которые можно будет применять в практике сельского хозяйства.
Биоразнообразие
эндофитных бактерий. Нишу эндофитов могут занять лишь те бактерии, которые
способны колонизировать межклеточные пространства различных частей растения, и
даже семян. Классические исследования биоразнообразия эндофитных бактерий
основывались на характеристике изолятов, полученных из внутренних тканей
различных растений после их тщательной поверхностной стерилизации.
Бактериальные эндофиты включают как грамположительные, так и грамотрицательные
виды, выделяемые из широкого спектра растений-хозяев.
1.3 Экологические
особенности тлей
Среди травянистых культурных растений наибольшие площади посевов в мире занимает пшеница. По существующим прогнозам ее потребление в составе продукции полевого растениеводства к 2020 году достигнет 40%.
Для многих государств пшеница представляет стратегический ресурс. В числе насекомых, вредящих пшенице, во всех районах ее возделывания отмечаются Настоящие тли. Они питаются как на листьях, так и на колосьях и корнях. Подавляющее большинство тлей имеют сложный жизненный цикл, включающий несколько партеногенетических поколений и одно полоносное, - у однодомных - на тех же травянистых растениях-прокормителях, а у двудомных, как правило, на деревьях или кустарниках.
У ряда видов с широкими ареалами, включающими несколько ландшафтно-климатических зон, в полупустынных и тропических регионах полоносное поколение может отсутствовать. Этими особенностями, в сочетании с составом фитоценозов обочин полей и прилежащих угодий, а также особенностями агротехники, определяются видовой состав тлей заселяющих полевые культуры, сроки и интенсивность их заселения и, в конечном счете, их потенциальная вредоносность.
Для лесостепной зоны России известно 23 вида тлей, питающихся на зерновых культурах.
Для развития и размножения тли большое значение имеют температурные условия. Для каждого вида характерен определённый температурный режим, при котором все жизненные процессы проходят наиболее интенсивно [7]. Большие отклонения от оптимума вызывают нередко гибель вредителя. Способность насекомых переносить длительное охлаждение различна не только у отдельных видов, но даже у одного вида в зависимости от его физиологического состояния. На размножение вредителей существенное влияние оказывает взаимосвязь насекомых с другими животными организмами [11]. В биоценозе складываются сложные «цепи питания», которые оказывают большое влияние на взаимоотношения компонентов, заселяющих определённый биотоп [14]. Например, различные виды тлей питаются соком растений, а выделяемые ими сахара служат пищей муравьям, наездникам и некоторым мухам. Тлями питаются многие виды хищных насекомых, например жуки и личинки кокцинеллид, личинки хризопы, личинки мух журчалок. Тлей и их врагов - хищных насекомых - поедают разнообразные насекомоядные птицы, которые, в свою очередь, становятся жертвами хищных птиц. Нарушение в какой-то части сложившихся «цепей питания» приводит в ряде случаев к существенному и непредвиденному или нежелательному изменению биоценоза в целом [13].
Различные сочетания факторов внешней среды вызывают у многих насекомых более или менее резкие изменения численности, причины которых у разных видов неодинаковы. Решающую роль могут играть наличие и состав пищи, погодные условия, воздействие хищников, паразитов и болезней и т. д. [11]. Колебания численности - следствие соотношений между плодовитостью вида и выживаемостью потомства, зависящей от условий среды и способности организма приспособиться к ним [14].
Отдельные виды тлей имеют свои
ареалы. Черемухово-злаковая тля встречается в СНГ: вся европейская часть, на север
до Хибин, Кавказ, Казахстан, Сибирь, Дальний Восток.; вязово-злаковая тля
отмечена на Кавказе, в Средней Азии, южном Приморье; ячменная тля - в степной и
лесостепной зонах европейской части, в Казахстане и Средней Азии. Обыкновенная
злаковая тля распространена в степной зоне, на Кавказе, в Средней Азии, на юге
Сибири. Большая злаковая тля встречается в СНГ: лесостепная и степная зоны
европейской части, Кавказ, юг Сибири и Дальний Восток, Средняя Азия [11].
1.4 Симбиотические
микроорганизмы тлей
Степень значимости симбионтов для
организма хозяина варьирует в зависимости от того, к какому классу он
относится. Значимость облигатных симбионтов значительно выше, чем
факультативных. К примеру, устойчивость тлей к высоким температурам, как
выяснилось, определяется генетическими особенностями их симбионтов - бактерий
Buchnera. Главная проблема, с которой сталкивается организм при перегревании -
это изменение вторичной структуры белков (денатурация). В большинстве случаев
клетка в ответ на повышение температуры активирует ряд генов, их называют
генами теплового шока - heat - shock genes, которые в свою очередь кодируют ряд
специфических белков, называемых шаперонами. Их функция заключается в том, что
они «насильно» придают белкам нативную конфигурацию, не позволяя им
денатурировать. Так же бактерии Buchnera играют ключевую роль в питании тли
[17]. Вместо тлей биосинтезом аминокислот занимаются бактерии-симбионты,
которые, в свою очередь, снабжают организм хозяина всем спектром аминокислот.
Установлено, что в число таких аминокислот входят аргинин, триптофан, треонин,
гистидин, лейцин, изолейцин, лизин, валин и фенилаланин. Бактерия синтезирует
гораздо больше аминокислот, чем это ей необходимо, и избыток аминокислот
достается насекомому. Так же известно, что симбионты обеспечивают тлю
необходимыми аминокислотами ещё на стадии эмбрионального развития.
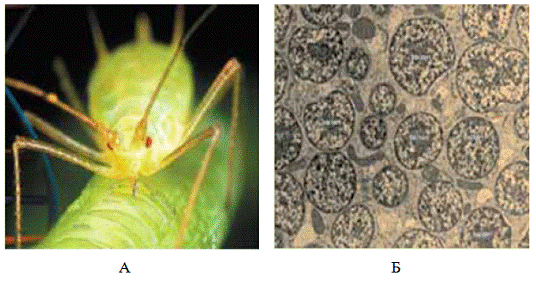
Рисунок 1 - Гороховая тля
Acyrthosiphon pisum Harris, 1776 (А) и внутриклеточный симбионт Buchnera (Б)
[17]
Установлено, что для нормального существования Buchnera в организме хозяина необходимо, чтобы насекомое получало достаточное количество калия, магния, железа, цинка, марганца, меди и кальция. Это было доказано экспериментально, путем выращивания тли на синтетической среде. Отсутствие вышеперечисленных элементов в рационе задерживало развитие симбионтов в первом или втором поколении хозяев и приводило к полной их гибели в последующих поколениях [18].является уникальным факультативным симбионтом, так как это единственный пример причастности эндосимбионтов к изменению окраски тела организма-хозяина. Наличие данного симбионта проявляется фенотипически. Именно он изменяет окраску красных тлей на зеленую. Доказано это было следующим образом: учеными была выведена линия красных тлей, для чистоты эксперимента с минимально отличающимися геномами. Данные насекомые были разделены на две опытные группы. Одну группу инфицировали бактериями Rickettsiella, а другую оставили в качестве контроля. В результате организмы, у которых присутствовали бактерии Rickettsiella изменили свой цвет. Это говорит о том, что Rickettsiella каким-то образом активизирует выработку зеленых пигментов, что вызывает изменение цвета тела [19].
Такая окраска тли в свою очередь делает её менее уязвимой для божьих коровок, но более уязвимой для атак наездников. Интересно, что большинство (около 80%) тлей, инфицированных Rickettsiella, были также инфицированы симбиотическими бактериями Hamiltonella или Serratia [20].