Материал: 1_2_Lektsia_Obmen_uglevodov_chast_2

Окисление цитоплазматического NADH+Н+ в ЦПЭ. Челночные системы
NADH + Н+ (иногда для простоты пишут просто NADH2) oбразующийся при окислении глицеральдегид-3-фосфата в аэробном гликолизе, подвергается окислению путѐм переноса атомов водорода в ЦПЭ. Однако митохондриальная мембрана для него непроницаема. Перенос водорода через мембрану происходит с помощью специальных «челночных» механизмов: водород транспортируется через мембрану при участии пар субстратов, связанных соответствующими дегидрогеназами, т.е., с обеих сторон митохондриальной мембраны находится специфическая дегидрогеназа.
Известны 2 челночных механизма для переноса водорода. В первой из них водород от NADH в цитозоле передаѐтся на ДГАФ ферментом глицерол-3- фосфатдегидрогеназой (NAD-зависимой). Образованный в ходе этой реакции глицерол-3-фосфат (другое название – α-глицерофосфат), окисляется далее ферментом внутренней мембраны митохондрий – глицерол-3- фосфатдегидрогеназой (FAD-зависимой). Затем протоны и электроны с FADH2 переходят на убихинон, и далее по ЦПЭ.
А-глицерофосфатный челночный механизм работает в клетках белых мышц и гепатоцитов. Однако в клетках сердечных мышц митохондриальная глицерол-3-фосфат-дегидрогеназа отсутствует.
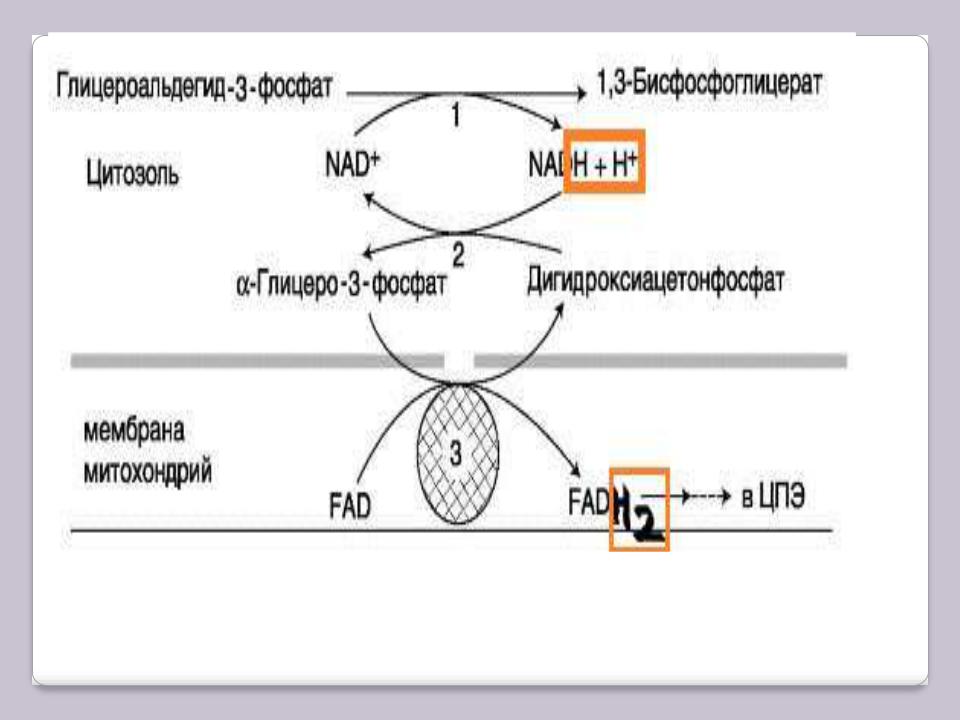
А-глицерофосфатный челночный механизм
1– глицеральдегид-3-фосфатдегидрогеназа;
2– глицерол-3-фосфатдегидрогеназа;
3– глицерол-3-фосфат-дегидрогеназа.

Малат-аспартатный челночный механизм, в
которой участвуют яблочная кислота (малат), цитозольная и митохондриальная малат-дегидрогеназы, является более универсальной.
В цитоплазме NADH восстанавливает щавелево-уксусную кислоту (оксалоацетат) в малат, который при участии переносчика проходит в митохондрии, где окисляется в оксалоацетат NAD-зависимой малатдегидрогеназой. Восстановленный в ходе этой реакции NAD отдаѐт водород в ЦПЭ. Однако образованный из малата оксалоацетат выйти самостоятельно из митохондрий в цитозоль не может, так как мембрана митохондрий для него непроницаема. Поэтому оксалоацетат путѐм трансаминирования* превращается в аспарагиновую кислоту (аспартат), который и транспортируется в цитозоль, где снова превращается в оксалоацетат.
Обе челночные системы отличаются по количеству синтезированного АТФ. В первой системе соотношение Р/O равно 2, так как водород вводится в ЦПЭ на уровне KoQ. Вторая система энергетически более эффективна, так как передаѐт водород в ЦПЭ через митохондриальный NAD+ и соотношение Р/О близко к 3.
* см. раздел «Азотистый обмен на II курсе»
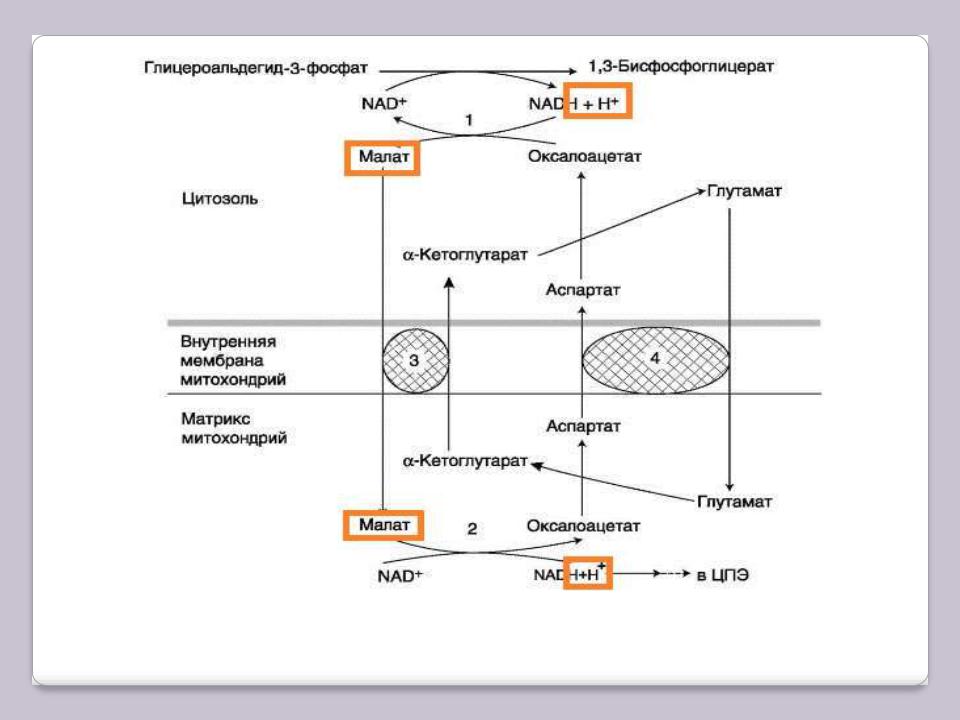
Малат-аспартатный челночный механизм
1, 2 – окислительно-восстановительны реакции, обеспечивающие транспорт водорода из цитозоля в митохондрии на ЦПЭ; 3, 4 – транслоказы, обеспечивающие транспорт α-кето-глутарата, аспартата и глутамата через мембрану митохондрий.
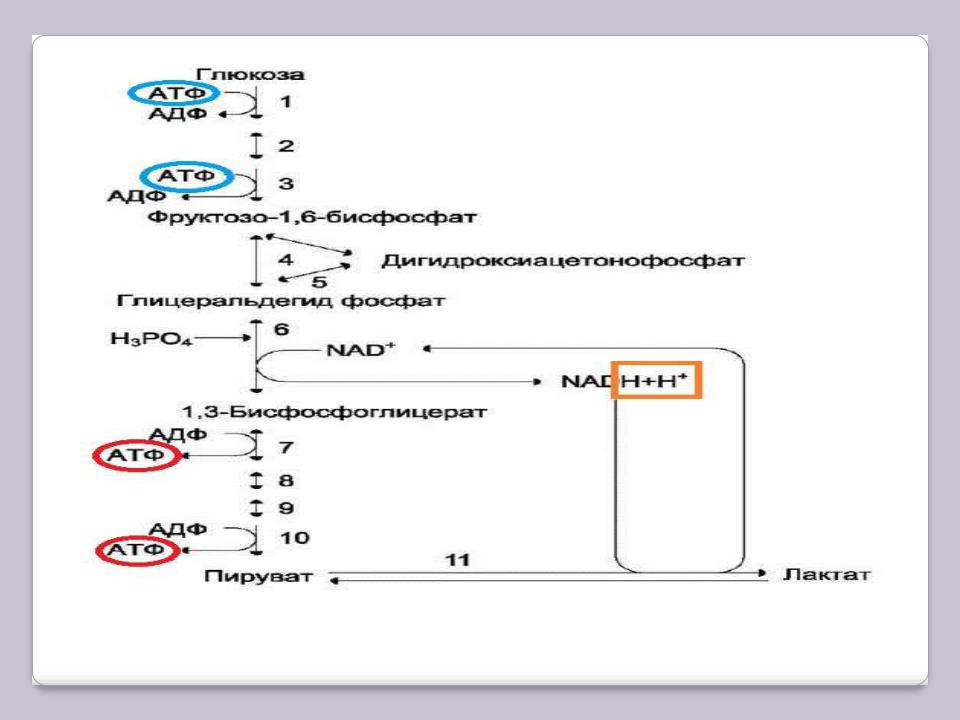
Анаэробный гликолиз
6, 11 – гликолитическая оксидоредукция.
Голубым цветом обведены затраты АТФ, красным – получение, оранжевым – водороды в составе восстановленных коферментов.