Материал: 1 курс Биология Экзамен Мед фак билеты
11 Билет
Рибонуклеиновые кислоты (РНК), тип нуклеиновых кислот, имеющих универсальное распространение в живой природе; содержат в качестве углеводного компонента рибозу, а в качестве азотистых оснований аденин и гуанин (пуриновые основания) и урацил и цитозин (пиримидиновые основания). В небольших количествах в состав Рибонуклеиновые кислоты входят также некоторые др. производные пурина и пиримидина. Рибонуклеиновые кислоты — линейные полинуклеотиды с длиной цепи от нескольких десятков до десятков тыс. нуклеотидов (молекулярная масса от 10—20×103 до 5—6×106), причём каждая индивидуальная Рибонуклеиновые кислоты имеет определённую последовательность нуклеотидов. В организме Рибонуклеиновые кислоты находятся главным образом в виде комплексов с белками — рибонуклеопротеидов. Рибонуклеиновые кислоты играют важнейшую биологическую роль во всех живых организмах, участвуя в реализации генетической информации и биосинтезе белков. Макромолекулярная структура Рибонуклеиновые кислоты представлена в основном однонитчатыми полинуклеотидными цепями, которые образуют двуспиральные участки по принципу комплементарности оснований. Многие вирусы содержат Рибонуклеиновые кислоты в качестве единственного нуклеинового компонента. В таких РНК-содержащих вирусах Рибонуклеиновые кислоты могут служить матрицей для биосинтеза не только РНК, но и ДНК (обратная транскрипция). Биосинтез Рибонуклеиновые кислоты осуществляется (главным образом в клеточном ядре) из рибонуклеозид-трифосфатов под действием ферментов полимераз на матрице дезоксирибонуклеиновых кислот (ДНК-зависимые РНК-полимеразы) или у некоторых вирусов на матрице Рибонуклеиновые кислоты (РНК-зависимые РНК-полимеразы). В клетках бактерий, животных и растений различные типы Рибонуклеиновые кислоты выполняют неодинаковые биологические функции и различаются по строению и метаболизму. Важнейшие типы РНК следующие. Рибосомные Рибонуклеиновые кислоты (рРНК) входят в состав рибосом и составляют основную массу клеточных Рибонуклеиновые кислоты; представлены Рибонуклеиновые кислоты с константами седиментации 23 S, 16 S и 5 S. Полностью установлена первичная структура (т. е. последовательность нуклеотидов) для 5 S РНК, почти полностью — для 16 S РНК и частично для 23 S РНК кишечной палочки. Размеры и структура рибосомных Рибонуклеиновые кислоты у организмов разных видов неодинаковы. Их биологическая роль не вполне выяснена, целостность их молекул необходима для биосинтеза белков в рибосомах. Транспортные Рибонуклеиновые кислоты (тРНК): имеют константу седиментации около 4 S и молекулярную массу около 25 000, т. е. являются сравнительно низкополимерными (около 80 нуклеотидных остатков); содержат относительно много метилированных и других минорных оснований. Их биологическая роль заключается в присоединении активированных аминокислотных остатков и переносе (транспорте) их на рибосомы, т. е. к месту синтеза полипептидных цепочек. Для каждой аминокислоты имеются свои специфические тРНК (обычно более одной). Все тРНК имеют сложную, частью двуспиральную, макромолекулярную структуру, изображаемую в виде клеверного листа. Они содержат участки, присоединяющиеся к рибосоме, триплет нуклеотидов (антикодон), присоединяющийся к кодону мРНК, и концевой участок, присоединяющий аминокислотный остаток. Первичная структура более 60 тРНК установлена полностью. Информационные, или матричные, Рибонуклеиновые кислоты (иРНК, или мРНК) представляют собой наиболее разнородную группу и играют роль матриц при биосинтезе белков в процессе трансляции (считывания нуклеотидного кода и перевода его в определённую последовательность аминокислот в полипептидных цепях белков). Все виды Рибонуклеиновые кислоты синтезируются в клетках на матрице ДНК, образуя последовательность рибонуклеотидов, комплементарную последовательности дезоксирибонуклеотидов в ДНК (процесс транскрипции). В клеточном ядре обнаружены гигантские молекулы — предшественники мРНК, большая часть которых распадается внутри ядра и только сравнительно небольшая часть молекулы переходит в цитоплазму и образует собственно мРНК. Быстро распадающаяся в клеточном ядре Рибонуклеиновые кислоты, вероятно, играет регуляторную роль.
РНК (рибонуклеиновая кислота), так же как и ДНК представляет собой полимер, мономерами которого служат нуклеотиды. Азотистые основания те же самые, что входят в состав ДНК (аденин, гуанин, цитозин), четвертое – урацил – присутствует в молекуле РНК вместо тимина. Нуклеотиды РНК содержат вместо дезоксирибозы другую пентозу – рибозу. В цепочке РНК нуклеотиды соединяются путем образования ковалентных связей между рибозой одного нуклеотида и остатком фосфорной кислоты другого.
Известны двух – и одно цепочечные молекулы РНК. Двухцепочечные РНК служат для хранения и воспроизведения наследственной информации у некоторых вирусов, т.е. у них выполняется функции хромосом. Одноцепочечные РНК осуществляют перенос информации о последовательности аминокислот в белках от хромосомы к месту их синтеза и участвуют в процессах синтеза.
Экспрессия генов – реализация информации, записанной в генах, осуществляемой в два этапа: транскрипция, трансляция.
Транскрипция – синтез РНК с использованием ДНК в качестве матрицы. В результате возникает 3 типа РНК: матричная (мРНК), рибосомная (рРНК), транспортная (тРНК).
Стадии транскрипции:
1). Инициация – образование нескольких начальных звеньев РНК.
2). Элонгация – продолжается дальнейшее расплетение ДНК и синтез РНК по кодирующей цепи.
Терминация – когда полимераза достигает терминатора (точки отсчета транскрипции), она немедленно отщепляется от ДНК, локальный гибрид ДНК-РНК разрушается и новосинтезированная РНК транспортируется из ядра в цитоплазму. Транскрипция заканчивается.
Оперон — функциональная единица генома у прокариот, в состав которой входят цистроны (гены, единицы транскрипции), кодирующие совместно или последовательно работающие белки и объединенные под одним (или несколькими) промоторами. Такая функциональная организация позволяет эффективнее регулировать экспрессию (транскрипцию) этих генов.
Начинается и заканчивается оперон регуляторными областями — промотором в начале и терминатором в конце, кроме этого, каждый отдельный цистрон может иметь в своей структуре собственный промотор и/или терминатор.
см. « 05.pdf » файл
Репрессия — подавление активности генов, чаще всего путём блокирования их транскрипции.
Индуктор — фактор (вещество, свет, теплота), вызывающий транскрипцию генов, находящихся в неактивном состоянии.
Эволюционное учение
Понадобился кропотливый гений великого шведского биолога Карла Линнея (1707-1778) для того, чтобы навести порядок в этих грудах материала, и создание Линнеем систематики как науки было первым и, пожалуй, самым важным камнем, заложенным в будущий фундамент дарвинизма. К. Линней был креационистом (он писал, что "видов столько, сколько их создало Бесконечное существо"). Однако его имя как ученого стоит рядом с такими именами, как И. Ньютон, Г. Лейбниц. Историческое значение К. Линнея состоит в том, что он выдвинул принцип иерархичности систематических категорий (таксонов): виды объединяются в роды, роды в семейства, семейства в отряды, отряды в классы и т.д. Иерархичность системы Линнея близка к идее расхождения признаков у родственных организмов, предложенной столетием позже Ч. Дарвином. К. Линней первым поместил человека среди отряда приматов, нанеся первый сокрушительный удар представлениям об исключительности человека как явления природы.
При этом Линней не утверждал, что человек произошел от обезьяны, он лишь подчеркнул несомненное внешнее сходство. Принцип иерархичности был сведен Линнеем в основном труде его жизни "Системе природы". Первое издание этой книги (1735 г.) имело лишь 13 страниц, десятое (1758 г.) - 1384 стр., а в двенадцатом (1778 г.) - 2335 стр. В течение всей жизни Линней продолжал работу по описанию новых видов и установлению иерархичности систематических категорий.
Принятие принципа иерархичности таксонов, стимулированное К. Линнеем бурное развитие систематики в большей степени способствовали возникновению и признанию дарвинизма, нежели труды трансформистов и ранних эволюционистов второй половины XVIII - первой половины XIX в.
Теория наследования приобретенных признаков Ламарка — эволюционная теория, созданная французским биологом Жаном Батистом Ламарком.
Всех животных Ламарк распределил по шести ступеням, уровням (или, как он говорил, «градациям») по сложности их организации. Дальше, совершенствуясь, организмы вынуждены приспосабливаться к условиям внешней среды. Как это происходит согласно теории Ламарка?
Для объяснения этого учёный сформулировал несколько «законов». Прежде всего, это «закон упражнения и не упражнения органов». Наибольшую известность из примеров, приведённых Ламарком, приобрёл пример с жирафами. Жирафам приходится постоянно вытягивать шею, чтобы дотянуться до листьев, растущих у них над головой. Поэтому их шеи становятся длиннее, вытягиваются. Муравьеду, чтобы ловить муравьёв в глубине муравейника, приходится постоянно вытягивать язык, и тот становится длинным и тонким. С другой стороны, кроту под землёй глаза только мешают, и они постепенно исчезают.
Если орган часто упражняется, он развивается. Если орган не упражняется, он постепенно отмирает.
Другой «закон» Ламарка — «закон наследования приобретённых признаков». Полезные признаки, приобретённые животным, по мнению Ламарка, передаются потомству. Жирафы передали потомкам вытянутую шею, муравьеды унаследовали длинный язык, и так далее.
Ч. Дарвину (1809 - 1882). Использовав весь накопленный к тому времени багаж знаний, он создал научно обоснованную теорию эволюци живой природы, названную в дальнейшем дарвинизмом.
В 1859 г. Ч. Дарвин опубликовал работу "Происхождение видов путем естественного отбора, или Сохранение благоприят-ствуемых пород в борьбе за жизнь", которая явилась результатом собственных наблюдений во время кругосветного путешествия на корабле "Бигль" и обобщения достижений науки и сельскохозяйственной практики того времени. Позже он написал такие работы, как "Изменение животных и растений под, влиянием одомашнивания" (1868), "Происхождение человека и половой отбор" (1871), "Действие перекрестного опыления и самоопыления в растительном мире" (1876) и др. В своих трудах Дарвин вскрыл механизм становления видов, т. е. механизм эволюции органического мира.
Дарвин считал, что все виды в природе способны размножиться в геометрической прогрессии. Это правило не знает исключений ни в растительном, ни в животном мире. Каждый вид способен произвести и производит гораздо больше особей, чем выживает их до взрослого состояния; юных особей всегда больше, чем взрослых. Однако число взрослых особей каждого вида растений и животных сохраняется более или менее постоянным. Исходя из этого он заключил, что часть особей гибнет в "борьбе за жизнь", в "борьбе за существование".
Основываясь на наблюдениях в природе, Дарвин обнаружил, что для растений и животных характерна всеобщая изменчивость признаков и свойств, так как даже в потомстве одной пары родителей нет совершенно одинаковых особей.
Синтетическая теория эволюции (СТЭ) — современная эволюционная теория, которая является синтезом различных дисциплин, прежде всего, генетики и дарвинизма. СТЭ также опирается на палеонтологию, систематику, молекулярную биологию и другие.
интетическая теория в её нынешнем виде образовалась в результате переосмысления ряда положений классического дарвинизма с позиций генетики начала XX века. После переоткрытия законов Менделя (в 1901 г.), доказательства дискретной природы наследственности и особенно после создания теоретической популяционной генетики трудами Рональда Фишера, Джона Б. С. Холдейна-младшего и Сьюэла Райта, учение Дарвина приобрело прочный генетический фундамент.
Статья С. С. Четверикова «О некоторых моментах эволюционного процесса с точки зрения современной генетики» (1926) по сути стала ядром будущей синтетической теории эволюции и основой для дальнейшего синтеза дарвинизма и генетики. В этой статье Четвериков показал совместимость принципов генетики с теорией естественного отбора и заложил основы эволюционной генетики. Главная эволюционная публикация С. С. Четверикова была переведена на английский язык в лаборатории Дж. Холдейна, но никогда не была опубликована за рубежом. В работах Дж. Холдейна, Н. В. Тимофеева-Ресовского и Ф. Г. Добржанского идеи, выраженные С. С. Четвериковым, распространились на Запад, где почти одновременно Р. Фишер высказал очень сходные взгляды о эволюции доминантности.
Толчок к развитию синтетической теории дала гипотеза о рецессивности новых генов. Говоря языком генетики второй половины XX века, эта гипотеза предполагала, что в каждой воспроизводящейся группе организмов во время созревания гамет в результате ошибок при репликации ДНК постоянно возникают мутации — новые варианты генов.
ТРИПАНОСОМЫ
А)Царство: Протисты
Тип: Эвгленовые
Класс: Кинетопластиды
Отряд: Трипаносоматиды
Семейство: Трипаносомовые
Б)Трипаносомы — род паразитических одноклеточных простейших семейства трипаносомовые, которые паразитируют на различных хозяевах и вызывают многие заболевания, среди которых сонная болезнь и болезнь Шагаса. Естественным резервуаром трипаносом в основном являются млекопитающие, переносчиком — насекомые.
Различные клеточные формы трипаносом
Для трипаносом характерно чередование клеточных форм: эпимастиготной в кишечнике насекомого и трипомастиготной и амастиготной в организме млекопитающих.
Различные клеточные формы трипаносом:
амастиготная форма — овальная или круглая, обычно встречается без жгутика
промастиготная форма — продолговатой формы, кинетопласт и кинетосома находятся в передней части клетки
эпимастиготная форма — продолговатой формы, кинетопласт и кинетосома находятся в задней части клетки
трипомастиготная форма — кинетопласт и кинетосома находятся сзади ядра, однако, в отличие от предыдущей формы, ундулирующая мембрана широкая и длинная
инвазивная или метацикличная форма — характерное отсутствие свободного жгутика.
Род трипаносом интересен наличием механизма защиты от иммунной системы жертвы. При проникновении в организм и их обнаружении иммунной системой у трипаносом включаются гены, ответственные за синтез гликопротеинов, в результате находящиеся на поверхности мембраны гликопротеины сменяются на другие, и иммунная система не может распознать паразитов, что дает им больше времени для размножения.
Размножаются трипаносомы делением надвое, хотя есть сведения о генетическом обмене между трофозоитами
БИЛЕТ №12
Реализация биологической информации в клетке.
Транскрипция - первая стадия реализации генетической информации в клетке. В ходе процесса образуются молекулы мРНК, служащие матрицей для синтеза белков, а также транспортные, рибосомальные и другие виды молекул РНК, выполняющие структурные, адапторные и каталитические функции
Транскрипция у эукариотов происходит в ядре. В основе механизма транскрипции лежит тот же структурный .принцип комплементарного спаривания оснований в молекуле РНК (G ≡ C, A=U и Т=А). ДНК служит только матрицей и в ходе транскрипции не изменяется. Рибонукле-озидтрифосфаты (ЦТФ, ГТФ, АТФ, УТФ) -субстраты и источники энергии, необходимые для протекания полимеразной реакции, образования 3',5'-фосфодиэфирной связи между рибонуклеозидмонофосфатами.
Синтез молекул РНК начинается в определённых последовательностях (сайтах) ДНК, которые называют промоторы, и завершается в терминирующих участках (сайты терминации). Участок ДНК, ограниченный промотором и сайтом терминации, представляет собой единицу транскрипции - транскриптон. У эукариотов в состав транскриптона, как правило, входит один ген у прокариотов несколько. В каждом транскриптоне присутствует неинформативная зона; она содержит специфические последовательности нуклеотидов, с которыми взаимодействуют регуляторные транскрипционные факторы.
Транскрипционые факторы - белки, взаимодействующие с определёнными регуляторными сайтами и ускоряющие или замедляющие процесс транскрипции. Соотношение информативной и неинформативной частей в транскриптонах эукариотов составляет в среднем 1:9 (у прокариотов 9:1).
Соседние транскриптоны могут быть отделены друг от друга нетранскрибируемыми участками ДНК. Разделение ДНК на множество транскриптонов позволяет осуществлять с разной активностью индивидуальное считывание (транскрипцию) разных генов.
В каждом транскриптоне транскрибируется только одна из двух цепей ДНК, которая называется матричной, вторая, комплементарная ей цепь, называется кодирующей. Синтез цепи РНК идёт от 5'- к З'-концу, при этом матричная цепь ДНК всегда антипараллельна синтезируемой нуклеиновой кислоте (рис. 4-28).
Транскрипция не связана с фазами клеточного цикла; она может ускоряться и замедляться в зависимости от потребности клетки или организма в определённом белке.
РНК-полимеразы
Биосинтез РНК осуществляется ДНК-зависимыми РНК-полимеразами. В ядрах эукариотов обнаружены 3 специализированные РНК-полимеразы: РНК-полимераза I, синтезирующая пре-рРНК; РНК-полимераза II, ответственная за синтез пре-мРНК; РНК-полимераза III, синтезирующая пре-тРНК. РНК-полимеразы - олигомерные ферменты, состоящие из нескольких субъединиц - 2α, β, β', σ. Субъединица о (сигма) выполняет регуляторную функцию, это один из факторов инициации транскрипции, РНК-полимеразы I, II, III, узнающие разные промоторы, содержат разные по строению субъединицы σ.
А. Стадии транскрипции
В процессе транскрипции различают 3 стадии: инициацию, элонгацию и терминацию.
Инициация
Активация промотора происходит с помощью большого белка - ТАТА-фактора, называемого так потому, что он взаимодействует со специфической последовательностью нуклеотидов промотора - ТАТААА- (ТАТА-бокс) (рис. 4-29).
Присоединение ТАТА-фактора облегчает взаимодействие промотора с РНК-полимеразой. Факторы инициации вызывают изменение кон-формации РНК-полимеразы и обеспечивают раскручивание примерно одного витка спирали ДНК, т.е. образуется транскрипционная вилка, в которой матрица доступна для инициации синтеза цепи РНК (рис. 4-30).
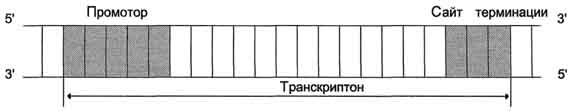
Рис. 4-27. Строение транскриптона.
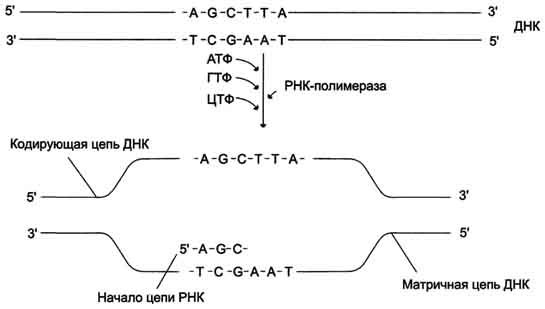
Рис. 4-28. Транскрипция РНК на матричный цепи ДНК. Синтез РНК всегда происходит в направлении 5' → 3'.
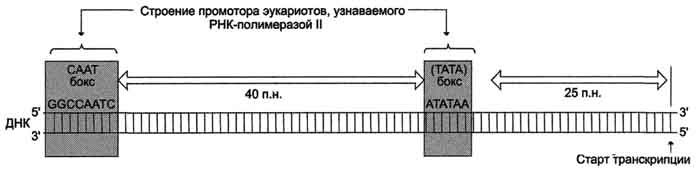
Рис. 4-29. Строение промотора эукариотов. Промоторные элементы - специфические последовательности нуклеотидов, характерные для любого промотора, связывающего РНК-полимеразу. Первый промоторный элемент - последовательность АТАТАА- (ТАТА-бокс) отделён от сайта начала транскрипции приблизительно на 25 пар нуклеотидов (п.н.). На расстоянии примерно 40 (иногда до 120) п.н. от него располагается последовательность GGCCAATC- (СААТ-бокс).
После того как синтезирован олигонуклеотид из 8-10 нуклеотидных остатков, σ-субъединица отделяется от РНК-полимеразы, а вместо неё к молекуле фермента присоединяются несколько факторов элонгации.
Элонгация
Факторы элонгации повышают активность РНК-полимеразы и облегчают расхождение цепей ДНК. Синтез молекулы РНК идёт от 5'- к З'-концу комплементарно матричной цепи ДНК. На стадии элонгации, в области транскрипционной
вилки, одновременно разделены примерно 18 нуклеотидных пар ДНК. Растущий конец цепи РНК образует временную гибридную спираль, около 12 пар нуклеотидных остатков, с матричной цепью ДНК. По мере продвижения РНК-полимеразы по матрице в направлении от 3'- к 5'-концу впереди неё происходит расхождение, а позади - восстановление двойной спирали ДНК.
Терминация
Раскручивание двойной спирали ДНК в области сайта терминации делает его доступным для фактора терминации. Завершается синтез РНК в
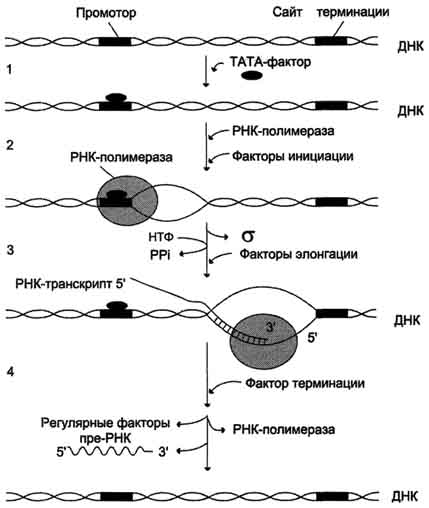
Рис. 4-30. Стадии транскрипции. 1 - присоединение ТАТА-фактора к промотору. Чтобы промотор был узнан РНК-полимера-зой, необходимо образование транскрипционного комплекса ТАТА-фактор/ТАТА-бокс (промотор). ТАТА-фактор остаётся связанным с ТАТА-боксом во время транскрипции, это облегчает использование промотора многими молекулами РНК-полимеразы; 2 - образование транскрипционной вилки; 3 - элонгация; 4.- терминация.