Материал: Інформаційні міжклітинні взаємодії. Клітинні рецептори та їхня участь у процесах міжклітинної сигналізації
Інформаційні міжклітинні взаємодії. Клітинні рецептори та їхня участь у процесах міжклітинної сигналізації
МІНІСТЕРСТВО ОСВІТИ І НАУКИ УКРАЇНИ
НАЦІОНАЛЬНИЙ ТЕХНІЧНИЙ УНІВЕРСИТЕТ УКРАЇНИ
«КИЇВСЬКИЙ ПОЛІТЕХНІЧНИЙ ІНСТИТУТ»
Факультет біотехнології і біотехніки
Кафедра
промислової біотехнології
РЕФЕРАТ
З дисципліни «Біологія клітини»
на
тему «Інформаційні міжклітинні взаємодії. Клітинні рецептори та їхня участь у
процесах міжклітинної сигналізації»
Виконав:
Студентка 1 курсу, група БТ-41
Хоруженко Наталія Костянтинівна
Перевірив:
Доцент каф. Промислово
біотехнології, к.б.н.
Жолнер Л.Г.
Київ
2014
ЗМІСТ
Вступ
. Типи міжклітинної регуляції
.1 Контактна регуляція
.2 Дистанційна (гуморальна) регуляція
. Передача сигналу
.1 Первинний і вторинний посередники
.2 Рецептори
.3 Характеристика молекулярних рецепторів і різних трансмітерів
Висновки
Список літератури
рецептор міжклітинний
синаптичний нейрон
ВСТУП
Багатоклітинний організм - це складна система, побудована із індивідуальних одиниць - клітин. В залежності від рівня еволюційного розвитку в одній живій істоті можуть поєднуватися десятки типів клітин, їхня кількість астрономічна, наприклад, людський організм в середньому складається зі 100 трильйонів (10014) клітин. З них формуються тканини, структури, органи, що, врешті, складають єдине ціле. Для функціонування такої надскладної системи окремі клітини мають взаємодіяти між собою, що зумовлює необхідність існування чіткої системи сигналізації.
Лише для підтримання гомеостазу
постійно відбувається порівняння «еталона» з інформацією, що прийшла від
організму. При виявленні розбіжності йде сигнал на коригування. Змінюється
«режим роботи». Знову порівняння, аналіз, сигнал на коригування.
Таблиця 1
Проте організм є динамічною системою і для напрямлених змін необхідна ще більш складна регуляція роботи його складових. Її основу складають міжклітинна комунікація та передача сигналу всередині окремих клітин, що дає можливість одній клітині контролювати поведінку інших. При цьому контроль може відбуватися як через прямий контакт між клітинами, так і дистанційно. Основу процесу можуть складати хімічні речовини та електричні імпульси, але варто зазначити, що і застосуванні імпульсів проходження сигналу через хімічний та мішаний синапси, яких більшість у організмі, базується на взаємодії хімічних речовин, на відміну від електричного синапса, в якому відбувається проходження імпульсу без затримки через прилеглі мембрани за рахунок конексонів. Хімічну міжклітинну сигналізацію класифікують за типом речовини, що бере участь у передачі сигналу: локальні хімічні медіатори, гормони і нейромедіатори.
Хоча на сьогоднішній день науці
багато відомо про комунікацію між клітинами, проте вивчення таких процесів
проблематичне. По-перше, вони відбуваються на молекулярному та атомному рівнях
та із високою швидкістю (їх тривалість - порядку мілісекунд в разі іонних
каналів і хвилин - у разі активації протеїнкіназ і ліпід-опосередкованих
кіназ). По-друге, вивчати такі взаємодії необхідно у живих клітинах, що залишає
відносно невеликий спектр методів дослідження. Найменші зміни у середовищі
спричиняють зміни метаболізму клітин, це спотворює результат і не дає
достовірної картини того, що відбувається у цілісному організмі. До другої
половини ХХ ст. уявлення про інформаційні взаємодії між клітинами і її
механізми базувалися на спостереженнях і теоретичних відомостях.
1. ТИПИ МІЖКЛІТИННОЇ РЕГУЛЯЦІЇ
.1 Контактна регуляція
При контактному способі регуляції регулююча і регульована клітини знаходяться один з одним в контакті.
Міжклітинні контакти мають таку класифікацію:
· Адгезійні, що механічно скріплюють клітини між собою. До них відносяться десмосоми (macula adherens), полудесмосоми і проміжний контакт (zonula adherens);
·
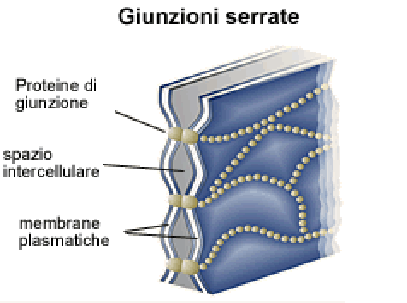
Рис. 1.1. Схема адгезійного зв’язку
Десмосомами об'єднуються дві структури. Одна з них (цитоплазматична платівка) здійснює зв'язок проміжних філаментів клітини з плазматичною мембраною; друга - зв'язок плазматичної мембрани з позаклітинним межмембранним матеріалом (десмоглея) в межах десмосоми. Десмосоми підтримують структурну цілісність тканини, скріплюючи клітини між собою. Десмосоми в комплексі з проміжними філаментами додають тканині пружність і підтримують у ній силу натягу. [1]
Проміжний контакт (оперізуюча
десмосома) - мембрани сусідніх клітин розділені проміжком шириною 10-20 нм,
заповненим аморфним або фібрилярним матеріалом. Електронно-щільна пластинка на
цитоплазматичній стороні клітинної мембрани в межах контакту містить білки
плакоглобін, вінкулін, α-актінін
і радіксін. До платівки вплетені кінці актинвмісних мікрофіламентів. В
утворенні контакту беруть участь трансмембранні білки адгезії з сімейства
кадгеринів. [2] Проміжний контакт не тільки скріплює мембрани
сусідніх клітин, але і стабілізує їх цитоскелет, об'єднуючи клітини з їх
вмістом в єдину жорстку систему подібно точковим адгезійним контактам
. 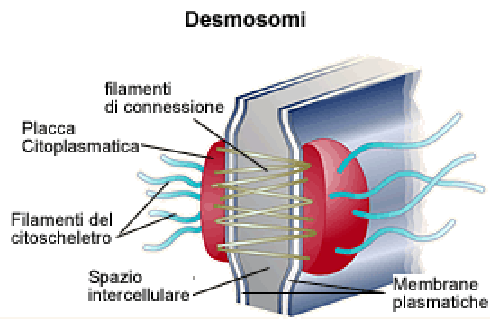
Рис. 1.2. Схема десмосоми
• Замикаючий, або щілинний, контакт (zonula occludens) формує в різних клітинних пластах бар'єр проникності, розмежовує різні за хімічним складом середовища;
Щілинний контакт забезпечує іонне і метаболічне сполучення клітин. Плазматичні мембрани клітин, утворюючих щілинний контакт, розділені щілиною шириною 2-4 нм. [1]
• Комунікаційні контакти. До них відносяться щілинні контакти і синапси.
У контактній регуляції важливу роль відіграє адгезія (здатність клітин вибірково прикріплюватися один до одного або до компонентів позаклітинного матриксу). Взаємодія трансмембранних молекул адгезії (в даному випадку - кадгеринів) забезпечує впізнавання клітинних партнерів і їх прикріплення один до одного, що дозволяє клітинам-партнерам формувати спеціалізовані міжклітинні контакти (наприклад, щілинні), а також передавати сигнали від клітини до клітини не тільки за допомогою дифундуючих молекул, але і шляхом взаємодії вбудованих в мембрану лігандів зі своїми рецепторами в мембрані клітини-партнера. [3]
Щілинні контакти, або нексуси (лат. Nexus), об'єднують окремі клітини в так звані функціональні сінцитії. Через щілинні контакти здійснюється електротонічна, метаболічна та інформаційна комунікація між контактуючими клітинами. Приклад функціонального сінцитія - міокард, як сукупність кардіоміоцитів. Головним елементом нексусу є не сама щелина, а наявність каналів з білків конексинів або інексинів, що регулярно її перетинають. Вони називаються конексонами. [2]
Конексон - трансмембранний білок циліндричної конфігурації, що складається з шести РЄ конексина. Два конексони сусідніх клітин з'єднуються в міжмембранному просторі і утворюють канал між клітинами. Канал конексона діаметром від 1,2 нм до 2,0 нм пропускає іони і молекули з M до 1,5 кД в обидві сторони. Відкритий чи закритий стан каналу конексона визначається конформацією конексинів, регулюється вмістом в цитозолі іонів Ca2+, pH, а також різницею електричного потенціалу пов'язаних клітин. [4]
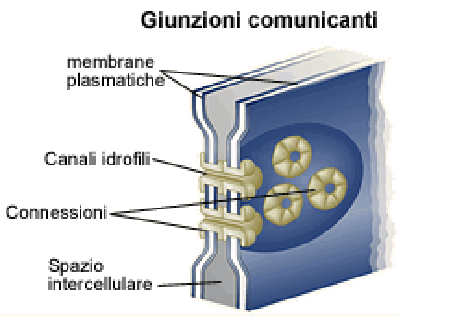
Рис. 1.3. Схема конексонів
Іншими словами, щілинні контакти забезпечують електричне сполучення пов'язаних клітин. Саме тому щілинні контакти забезпечують поширення збудження - миттєвий перехід іонів між клітинами.
Класичний приклад контактної регуляції - нервова регуляція. Нервова регуляція здійснюється за допомогою синапсів - між регулюючої і регульованою клітинами. Передачу сигналу в синапсі здійснюють спеціальні молекули, що звільняються з пресинаптичної клітини (нейромедіатори), а реєстрація сигналу відбувається за допомогою молекулярних рецепторів, вбудованих в постсинаптичну мембрану (так, в нервово-м'язовому синапсі між термінальними розгалуженнями α-мотонейронів і скелетними м'язовими волокнами діє ацетилхолін, що зв'язується з нікотиновими ацетилхоліновими рецепторами).
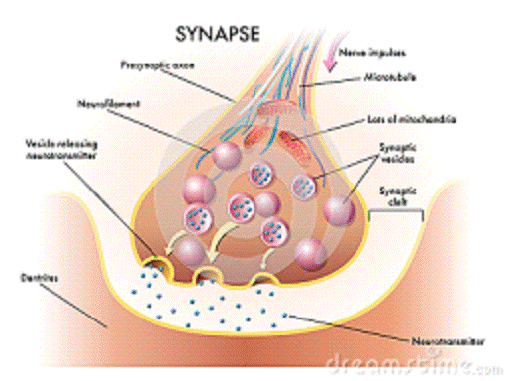 Рис. 1.4. Будова
синапса
Рис. 1.4. Будова
синапса
.2 Дистанційна (гуморальна)
регуляція
Дистанційна (гуморальна) регуляція характеризується тим, що регулююча і регульована клітини розташовані на відстані один від одного, іноді на досить значному. Для дистантной регуляції необхідний матеріальний агент передачі сигналу між дистантно розташованими клітинами. Такими агентами є інформаційно активні речовини (гормони, цитокіни, фактори росту і т.д.). Молекули таких речовин на шляху від клітини-регулятора до регульованої клітини-мішені знаходяться в різних рідинах (лат. Humor) організму: в крові, лікворі, лімфі, тканинній рідині. [5]
Приклад гуморальної регуляції - ендокринна регуляція: гормони, секретуються ендокринними клітинами, виділяються в рідини організму (у більшості випадків - в кров), рідиною транспортуються до клітин-мішеней, зв'язуються зі специфічними саме для конкретного гормону молекулярними рецепторами і змінюють режим функціонування клітини-мішені.
Гормони циркулюють в крові або вільно, або в комплексі зі зв'язуючими їх білками (T4, Т3, стероїдні гормони, інсуліноподібний фактор росту, гормон росту). Зв'язування з такими білками значно збільшує час напівжиття гормонів. Так, T4 в складі комплексу циркулює близько одного тижня, тоді як час напівжиття вільного T4 становить кілька хвилин.
Існує два варіанти дистанційної
регуляції: паракринна регуляція - ефект гормону, що впливає шляхом
дифузії на клітини-мішені; аутокринна регуляція - вплив гормону
безпосередньо на секретуючу цей гормон клітину. [6]
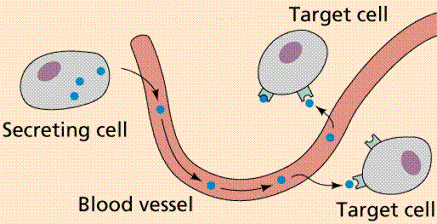
Рис. 1.5. Гуморальна регуляція
2. ПЕРЕДАЧА СИГНАЛУ
В узагальненому вигляді передача сигналу відбувається за наступною схемою:
. Взаємодія зовнішнього агента (стимулу, первинного посередника) з клітинним рецептором.
. Активація ефекторної молекули, що знаходиться в мембрані і відповідає за генерацію вторинних посередників.
. Утворення вторинних посередників.
. Зникнення посередника.
[2]
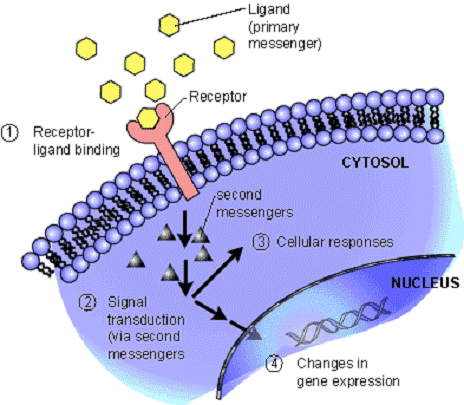
Рис. 2.1. Загальна схема передачі сигналу
.1 Первинний і вторинний посередники
Первинні посередники
Первинні посередники - це хімічні сполуки або фізичні фактори (квант світла), здатні активувати механізм передачі сигналу в клітині. Відносно клітини-мішені первинні посередники є екстраклітинними сигналами. В якості екстраклітинного стимула можуть виступати і молекули, що в достатку присутні всередині клітини, але в нормі в дуже низькій концентрації в міжклітинному просторі (наприклад, АТФ або глутамат). [3] Залежно від функцій первинні посередники можуть бути розділені на декілька груп:
· Гормони.
Гормон - це біологічно-активна
хімічна речовина, що виділяється ендокринними залозами безпосередньо у кров і
впливає на певні органи і тканини-мішені або на організм в цілому.
Таблиця 2.1
Т
Біологічний результат дії кожного гормону вельми специфічний. Хоча у клітині-мішені гормони змінюють зазвичай менше 1% білків і РНК, цього виявляється цілком достатньо для отримання відповідного фізіологічного ефекту. Більшість гормонів характеризуються трьома особливостями: вони розчиняються у воді; не зв'язуються з білками носіями; починають гормональний процес, як тільки з'єднуються з рецептором, який може знаходитися в ядрі клітки, її цитоплазмі, або розташовуватися на поверхні плазматичної мембрани. Найбільш важливі посередники для таких гормонів - цАМФ(циклічний аденозинмонофосфат), інозитолтрифосфат, іони кальцію. Так, в середовищі, позбавленому іонів кальцію, або в клітинах з недостатньою їх кількістю, дія багатьох гормонів послаблюється; при застосуванні речовин, що збільшують внутрішньоклітинну концентрацію кальцію, виникають ефекти, ідентичні до дії деяких гормонів.
· Цитокіни.
Цитокіни - це невеликі пептидні
інформаційні молекули. Виділяються на поверхню клітини А і взаємодіють з
рецептором клітини В, що знаходиться поруч. Таким чином, від клітини А до
клітини В передається сигнал, який запускає в клітині В подальші реакції.
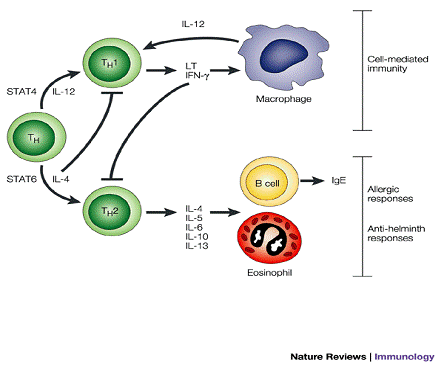
Рис. 2.2. Схема реалізації цитокінів
Їх основними продуцентами є лімфоцити. Крім лімфоцитів їх секретують макрофаги, гранулоцити, ретикулярні фібробласти, ендотеліальні клітини та інші типи клітин. Вони регулюють міжклітинні і міжсистемні взаємодії, визначають виживаність клітин, стимуляцію або пригнічення їх росту, диференціацію, функціональну активність і апоптоз, а також забезпечують узгодженість дії імунної, ендокринної та нервової систем в нормальних умовах і у відповідь на патологічний вплив. [7]
· Нейротрансмітери (нейромедіатори).
Біологічно активні хімічні речовини, за допомогою яких здійснюється передача електричного імпульсу від нервової клітини через синаптичний простір між нейронами, а також, наприклад, від нейронів до м'язової тканини. Нервовий імпульс, що надходить в пресинаптичного закінчення, викликає звільнення в синаптичну щілину медіатора. Молекули медіаторів реагують зі специфічними рецепторними білками клітинної мембрани, ініціюючи ланцюг біохімічних реакцій, що викликають зміну трансмембранного струму іонів, що призводить до деполяризації мембрани і виникнення потенціалу дії. Традиційно нейромедіатори відносять до трьох груп: амінокислоти, пептиди, моноаміни (у тому числі катехоламіни). В пресинаптичнії клітині везикули, що містять нейромедіатор, вивільняють його локально в дуже малий об’єм синаптичної щілини. Вивільнений нейромедіатор потім дифундує через щілину і зв'язується з рецепторами на постсинаптичні мембрані. Дифузія є повільним процесом, але перетин такої короткій дистанції, яка розділяє при- і постсинаптичну мембрани (0,1 мкм або менше), відбувається досить швидко і дозволяє здійснювати швидку передачу сигналу між нейронами або між нейроном і м'язом. [8]
· Фактори росту.
Клас невеликих природних пептидів та білків, що беруть участь в сигнальних системах організму еукаріотів, зв'язуючись із рецепторами на поверхні клітин із головною метою стимулювання їх росту та диференціації. Важливі для регулювання різноманітності клітинних процесів. Конкретний ефект на клітину та тип клітин, на які діє фактор росту, залежить від конкретного фактору. Наприклад, білок морфогенезу кісток стимулює диференціацію клітин кісток, тоді як фактор росту фібробластів і васкулоендотеліальний фактор росту стимулюють диференціювання кровоносних судин (ангіогенез). [7] На відміну від гормонів, фактори росту секретуються локально та мають обмежену область дії, тоді як гормони переносяться кровотоком і мають ефект на дуже віддаленні тканини організму.
Вторинні посередники
У відповідь на приєднання ліганду чи фізичний стимул багато мембранних рецепторів ініціюють синтез або деградацію внутрішньоклітинних малих молекул, що зв'язуються із цитоплазматичними рецепторами. Внутрішньоклітинні ліганди називають вторинними посередниками або месенджерами (на відміну від первинних посередників - позаклітинних).