Материал: В-Л-Быков-Цитогогия_и_общая_гистология
сверху покрыта уплощенными леммоцитамн и базальной мембраной, переходящей с мышечного волокна. В терминалях аксона имеются митохондрии и синаптические пузырьки, содержащие ацетилхолин.
Синаптическая щель шириной около 50 нм располагается между плазмолеммой ветвлений аксона и мышечного волокна; она содержит материал базальной мембраны и отростки глиальных клеток, разделяющих соседние активные зоны одного окончания.
Постсинаптическая часть представлена мембраной мышечного волокна (сарколеммой), образующей многочисленные складки (вторичные синаптические щели), которые увеличивают общую площадь щели и заполнены материалом, являющимся продолжением базальной мембраны. В области нервно-мышечного окончания мышечное волокно не имеет исчерченности, содержит многочисленные митохондрии, цистерны грЭПС, рибосомы и скопления ядер.
Рис. 14-13. Нервно-мышечное окончание (по Rohen J.W , Lutjen-Drecoll E. 1982). 1 - общий вид, 2 - ультраструктурная организация Нервно-мышечное окончание (НМО) состоит из пресинаптической части, образованной концевым ветвлением аксона (А), постсинаптической части - специализированного участка на мышечном волокне (MB) и разделяющей их первичной синаптической щели (ПСЩ). Нервное волокно (НВ) у MB утрачивает миелиновую оболочку и отдает веточки, сверху покрытые леммоцитами (ЛЦ) и базальной мембраной. Терминали А содержат митохондрии (МТХ) и синаптические пузырьки (СП). Мембрана мышечного волокна - сарколемма - образует многочисленные складки (СС), вследствие чего формируются вторичные синаптические щели (ВСЩ). В области НМО исчерченность MB отсутствует, здесь содержатся многочисленные МТХ, цистерны грЭПС, рибосомы и скопления ядер (Я). МФ - миофибриллы, ЯЛЦ - ядро ЛЦ.
- 481 -
Механизм передачи нервного импульса на мышечное волокно в нервно-
мышечном синапсе сходен с таковым в химическом межнейронном синапсе. При деполяризации пресинаптической мембраны происходит выделение ацетилхолина в синаптическую щель; его связывание с холинорецепторами в постсинаптической мембране вызывает ее деполяризацию и последующее сокращение мышечного волокна. Медиатор отщепляется от рецептора и быстро разрушается ферментом ацетилхолинэстеразой, который содержится в синаптической щели.
Понимание механизмов передачи возбуждения в нервно-мышечных окончаниях имеет существенное клиническое значение. Действие некоторых ядов (например, кураре) обусловлено блокированием этой передачи, вызванным их прочным связыванием с холинорецепторами. Получены аналоги таких веществ (миорелаксанты), которые нашли применение в хирургии для расслабления мышц при полостных операциях, проводимых в условиях искусственного дыхания (поскольку нарушается деятельность дыхательных мышц). При миастении (myasthenia gravis) – заболевании, которое характеризуется прогрессирующей мышечной слабостью, в сарколемме снижается содержание рецепторов ацетилхолнна (по-видимому, вследствие аутоиммунного процесса).
Двигательные нервные окончания в сердечной и гладких мышцах
имеют вид варикозно расширенных (до 0.5-2 мкм) участков тонких (0.1-0.5 мкм) веточек аксонов, которые содержат многочисленные синаптические пузырьки и митохондрии. Обычно они отделены от мышечных клеток широкой (около 100 нм) щелью. Как правило, иннервированы лишь отдельные клетки, возбуждение с которых передается на соседние посредством щелевых соединений.
Секреторные нервные окончания представляют собой конечные участки тонких аксонных веточек. Одни из них, утрачивая оболочку из леммоцитов, проникают сквозь базальную мембрану и располагаются между секреторными клетками, заканчиваясь терминальными варикозными расширениями, содержащими пузырьки и митохондрии (гиполеммальный нейроэффекторный контакт). Другие не проникают сквозь базальную мембрану, образуя варикозные расширения вблизи секреторных клеток (эпилеммальный нейроэффекторный контакт). Секреторные нервные окончания оказывают на железистые клетки несколько видов воздействия: гидрокинетическое (мобилизация воды), протеокинетическое (секреция белка), синтетическое (усиление синтеза) и трофическое (поддержание нормальной структуры и функции).
- 482 -
РЕЦЕПТОРНЫЕ (ЧУВСТВИТЕЛЬНЫЕ) НЕРВНЫЕ ОКОНЧАНИЯ
Рецепторные (чувствительные) нервные окончания воспринимают сигналы из внешней среды (экстерорецепторы) и внутренних органов (интерорецепторы). В зависимости от природы раздражения, регистрируемого рецепторами, они подразделяются в соответствии с физиологической классификацией, на механорецепторы, хеморецепторы, терморецепторы и болевые рецепторы (ноцицепторы). В специализированных органах чувств (орган вкуса, обоняния, зрения, равновесия и слуха) имеются особые рецепторные клетки, которые воспринимают соответствующие раздражения.
Морфологическая классификация чувствительных нервных окончаний
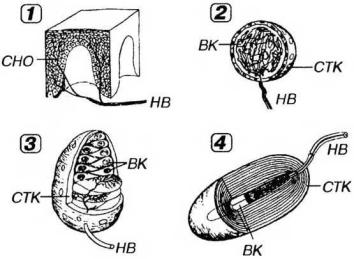
основана на особенностях их структурной организации. В соответствии с этой классификацией различают свободные и несвободные чувствительные нервные окончания; последние включают инкапсулированные и неинкапсулированные окончания (рис. 14-14).
Рис. 14-14. Рецепторные (чувствительные) нервные окончания (по Rohen J.W., Lutjen-Drecoll E. 1982. с изменениями). 1 - свободные нервные окончания (СНО) образованы терминальными ветвлениями дендрита чувствительного нейрона, идущего в составе нервного волокна (НВ). Несвободные инкапсулированные нервные окончания (2-4) образованы ветвлениями дендрита, окруженными леммоцитами, в совокупности с которыми они образуют структуру, называемую внутренней колбой (ВК). Снаружи окончания покрыты соединительнотканной капсулой (СТК). 2 - колба Краузе, 3 - осязательное тельце (Мейснера), 4 - пластинчатое тельце (Фатер-Пачини).
- 483 -
Свободные чувствительные нервные окончания состоят только из терминальных ветвлений дендрита чувствительного нейрона. Они встречаются в эпителии, а также в соединительной ткани. Проникая в эпителиальный пласт, нервные волокна утрачивают миелиновую оболочку и нейролемму, а базальная мембрана их леммоцитов сливается с эпителиальной. Свободные нервные окончания обеспечивают восприятие температурных (тепловых и холодовых), механических и болевых сигналов.
Несвободные чувствительные нервные окончания содержат все компоненты нервного волокна. Они разделяются на инкапсулированные (имеющие особую соединительнотканную капсулу) и неинкапсулированные.
Несвободные неинкапсулированные нервные окончания состоят из ветвлений дендритов, окруженных леммоцитами. Они встречаются в соединительной ткани кожи (дерме), а также собственной пластинки слизистых оболочек.
Несвободные инкапсулированные нервные окончания весьма разнообразны, но имеют единый общий план строения: их основу составляют ветвления дендрита, которые непосредственно окружены леммоцитами и снаружи покрыты особой соединительнотканной капсулой (см. рис. 14-14). К этому виду нервных окончаний относят пластинчатые тельца (Фатер-Пачини), осязательные тельца (Мейснера) тельца Руффини, колбы Краузе, нервномышечные веретена и нервно-сухожильные веретена (сухожильные органы Гольджи) .
Пластинчатые тельца (Фатер-Пачини) встречаются в соединительной ткани внутренних органов и кожи. Они имеют вид округлых образований диаметром 1-5 мм, воспринимают давление и вибрацию. Структурными компонентами тельца являются:
1)внутренняя колба (луковица), образованная видоизмененными уплощенными леммоцитами, в которую проникают одно или несколько нервных волокон, имеющих прямой ход;
2)наружная колба - слоистая соединительнотканная капсула, состоящая из фибробластов и коллагеновых волокон, образующих 10-60 концентрических пластин, между которыми имеется жидкость.
При деформации пластин капсулы давление передается на нервное окончание, что вызывает деполяризацию его мембраны.
Осязательные тельца (Мейснера) расположены преимущественно в сосочковом слое дермы, имеют эллипсоидную форму и небольшие
- 484 -
размеры (около 50-120 мкм). Их внутренняя колба состоит из плоских глиальных клеток, лежащих перпендикулярно длинной оси тельца, между которыми располагаются веточки дендритов. Между глиальными клетками проникают коллагеновые фибриллы, связанные с базальным слоем эпителия. Капсула тонкая, переходит в периневрий.
Тельца Руффини лежат в соединительнотканной части кожи и капсулах суставов; они воспринимают давление и имеют вид веретеновидных структур длиной до 1-2 мм. Внутреннюю колбу образуют глиальные клетки, между которыми располагаются многочисленные ветвящиеся терминали дендритов с расширениями на концах. Капсула хорошо выражена, образована коллагеновыми волокнами.
Колбы Краузе - мелкие (40-150 мкм) округлые тельца, являющиеся механорецепторами и, возможно, холодовыми рецепторами. Они расположены в соединительной ткани сосочкового слоя дермы и собственной пластинке слизистой оболочки полости рта, надгортанника, в конъюнктиве глаза. Внутренняя колба образована уплощенными глиальными клетками, между которыми тонкие веточки дендрита образуют сплетение в виде клубочка. Капсула состоит из плоских клеток, являющихся продолжением периневрия.
Нервно-мышечные веретена - рецепторы растяжения волокон поперечнополосатых мышц - сложные инкапсулированные нервные окончания, обладающие как чувствительной, так и двигательной иннервацией. Число веретен в мышце зависит от ее функции и тем выше, чем более точными движениями она обладает. Нервно-мышечное веретено (рис. 14-15) имеет длину 0,5-7 мм и располагается параллельно ходу волокон мышцы, называемых экстрафузальными (от лат. extra - вне и fuso - веретено, т.е. расположенными за пределами веретена). Веретено покрыто тонкой соединительнотканной капсулой (продолжением периневрия), внутри которой находятся тонкие поперечнополосатые штрафу зальные мышечные волокна двух видов:
-волокна с ядерной сумкой - в расширенной центральной части которых содержатся скопления ядер (1-4 волокна/веретено);
-волокна с ядерной цепочкой - более тонкие с расположением ядер в виде цепочки в центральной части (до 10 волокон/веретено).
Чувствительные нервные волокна образуют кольцеспиральные окончания на центральной части интрафузальных волокон обоих типов и гроздьевидные окончания у краев волокон с ядерной цепочкой.
Двигательные нервные волокна - тонкие, образуют мелкие нервномышечные синапсы по краям интрафузальных волокон, обеспечивая их тонус.
- 485 -