Материал: В-Л-Быков-Цитогогия_и_общая_гистология
Анемия (от греч аn - отсутствие, haima - кровь) - снижение содержания гемоглобина в крови при падении его уровня в отдельном эритроците и (или) концентрации эритроцитов в крови. Она может вызываться нарушением синтеза гемоглобина (вследствие недостаточности железа или образования его аномальных форм), кровопотерей, чрезмерным разрушением эритроцитов или их недостаточным образованием. При анемии страдают все системы организма, в первую очередь, вследствие недостаточного поступления кислорода в ткани.
Полицитемия (эритроцитоз) - повышение концентрации эритроцитов - может быть проявлением реакции адаптации, например, у люей, живущих на больших высотах (при низком содержании кислорода в воздухе). Полицитемия опасна из-за повышения вязкости крови, которое может приводить к нарушениям ее циркуляции.
Скорость оседания эритроцитов (СОЭ). При помещении крови в пробирку и предотвращении ее свертывания эритроциты формируют агрегаты в виде монетных столбиков и постепенно оседают на дно. Скорость оседания эритроцитов (СОЭ) зависит от многих факторов и в среднем выше у женщин, чем у мужчин. В норме она равна 5-9 мм/ч (по другим данным - 2-12 мм/ч). Этот показатель определяется при анализе крови и имеет существенное диагностическое значение, поскольку он резко увеличивается при многих инфекционных, воспалительных и онкологических заболеваниях.
Строение эритроцитов
Форма эритроцитов - двояковогнутый диск (рис. 7-2) - определяет более светлую окраску их центральной части по сравнению с периферической. Благодаря такой форме обеспечиваются;
(1)увеличение их поверхности (общая ее площадь составляет у взрослого около 3800 м2, что в 2000 раз превосходит поверхность тела); площадь поверхности каждого эритроцита примерно в 1.5 раза больше, чем у сферы такого же объема;
(2)снижение диффузионного расстояния (между поверхностью и наиболее удаленной от нее части цитоплазмы) - на 30% по сравнению с такими же элементами сферической формы, благодаря чему создаются оптимальные условия для газообмена;
(3)возможность увеличения объема эритроцита без повреждения его плазмолеммы благодаря наличию ее резерва, в частности, способность набухать
вгипотоничной среде;
(4)способность к обратимой деформации при прохождении через узкие и изогнутые капилляры.
-166 -
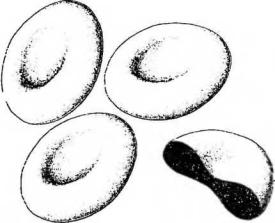
Рис. 7-2. Эритроциты: трехмерное изображение в СЭМ. На разрезе видна цитоплазма эритроцитов, обладающая высокой электронной плотностью.
Поддержание формы эритроцитов обеспечивается вследствие осмотического равновесия, которое достигается благодаря деятельности ионных насосов в их плазмолемме, а также особыми элементами цитоскелета (см. ниже).
Изменения формы эритроцитов возникают при их старении и в патологических условиях вследствие нарушений осмотического равновесия или (и) дефектов цитоскелета. В частности, сферическая форма эритроцитов, наблюдаемая при врожденном сфероцитозе, сопровождается их неспособностью к растяжению, деформации, осмотической нестойкостью и усиленным разрушением. Форма эритроцитов может изменяться также при образовании патологических форм гемоглобина. Так, точечная мутация гена, связанная с замещением одной аминокислоты в молекуле нормального гемоглобина взрослого человека (НЬА), приводит к появлению гемоглобина Б (НЬБ), который, теряя кислород, подвергается полимеризации с образованием агрегатов, механически деформирующих эритроциты (возможно, вследствие взаимодействия с элементами цитоскелета, связанными с плазмолеммой). Такие эритроциты, приобретающие серповидную форму, характеризуются малой гибкостью и сниженной продолжительностью жизни, свойственной серповиднокле-точной анемии
.
Пойкилоцитоз (от греч. роikilos - разнообразный и cytos, kytos, - клетка) - наличие в крови эритроцитов необычной формы.
- 167 -
Размеры эритроцитов: средний диаметр составляет 1.2-1.5 мкм (с отклонениями в обе стороны для большинства не более 0.5 мкм), толщина в краевой зоне - 1.9-2.5 мкм, в центральной - 1 мкм. По мере старения эритроцитов их размеры несколько уменьшаются.
Макроциты - (от греч. makros - большой, cytos, или kytos - клетка) - крупные эритроциты (с диаметром свыше 9 мкм), их преобладание в мазке крови называется макроцитозом.
Микроциты - (от греч. mikros - мелкий, cytos, или kytos - клетка) - мелкие эритроциты (с диаметром б мкм и менее), их повышенное содержание в мазке именуется микроцитозом.
Анизоцитоз (от греч. аn - отрицание, iso - равный, cytos, или kytos - клетка) - резкие различия в размерах отдельных эритроцитов на мазке.
Плазмолемма эритроцитов является самой толстой (20 нм) и наиболее изученной мембраной из всех биологических мембран. Она содержит рецепторы иммуноглобулинов, компонентов комплемента и ряда других веществ. В ее состав входят многочисленные интегральные и периферические белки, участвующие в транспортных процессах (в качестве ионных насосов, каналов, переносчиков) и обеспечивающие прикрепление элементов цитоскелета. Она обладает гибкостью, прочностью, растяжимостью, резистентностью к окислению, протеолизу и влиянию других повреждающих факторов. На наружной поверхности плазмолемма эритроцитов несет антигены Rh и детерминанты групп крови.
Цитоплазма эритроцитов оксифильна и обладает высокой электронной плотностью (см. рис. 7-2); органеллы в ней отсутствуют, могут встречаться лишь единичные мембранные пузырьки. Она содержит 66% воды, гемоглобин в виде гранул диаметром 4-5 нм, глюкозу, АТФ, ряд ферментов. Основной источник энергии эритроцитов - анаэробный гликолиз.
Цитоскелет эритроцитов образован рядом периферических и трансмембранных белков (рис. 7-3). В его состав входят: спектрин, гликофорин, анкирин, белки полосы 3 и полосы 4.1. Последние два названия отражают положение фракций при электрофорезе белков мембраны эритроцита. Белок полосы 3 выполняет помимо цитоскелетных функций роль анионного транспортного белка, обеспечивающего процессы газообмена.
Спектрин - периферический белок, служащий главным элементом цитоскелета эритроцита. Его молекула состоит из двух перекрученных
- 168 -
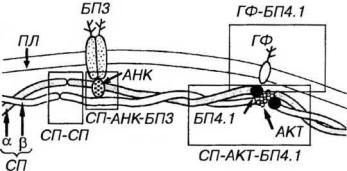
цепей - димеров (α- и β), которые стыкуются друг с другом "конец в конец". Он образует гибкую двумерную сеть филаментов на внутренней поверхности плазмолеммы эритроцита. Эти филаменты связаны в узлы с помощью актина и белка полосы 4.1 к прикреплены к трансмембранному белку полосы 3 посредством анкирина. Белок полосы 4.1 может связываться с цитоплазматическим доменом другого трансмембранного белка - гликофорина. В состоянии покоя спектриновые цепи скручены; при деформации в одних участках они распрямляются и вытягиваются, в других - скручиваются еще сильнее, благодаря чему происходит изменение формы эритроцита без изменения площади его поверхности. При более значительной деформации, требующей увеличения поверхности, может нарушиться связь элементов цитоскелета с плазмолеммой, и возникшая деформация станет необратимой или произойдет фрагментация плазмолеммы.
Рис. 7-3. Цитоскелет эритроцитов. Трансмембранные белки - белок полосы 3 (БПЗ) и гликофорин (ГФ) - пронизывают липидный бислой плазмолеммы (ПЯ). Димеры спектрина (СП) - а-СП и р-СП - стыкуются друг с другом "конец в конец”, образуя двумерную сеть филаментов на внутренней поверхности ПЛ эритроцита. Их прикрепление к ПЛ опосредуется анкирином (АНК), связывающим СП с БПЗ, а также комплексами, содержащими актин (АКТ) и белок полосы 4.1 (БП4.1), которые прикрепляют СП к ГФ. Варианты механизмов взаимодействия белков цитоскелета между собой и с мембранными белками выделены рамками.
Благодаря описанному устройству цитоскелета эритроцит обладает гибкостью и способен обратимо деформироваться в мелких сосудах. При врожденном сфероцитозе (см. выше) и некоторых других заболеваниях изменения формы и свойств эритроцитов обусловлены дефицитом спектрина, анкирина и нарушением связывания спектрина с другими белками цитоскелета эритроцита.
- 169 -
Ретикулоциты - молодые формы эритроцитов, недавно поступившие в кровоток из костного мозга. В них сохраняются митохондрии, небольшое число рибосом, центриоль и остатки комплекса Гольджи; ЭПС отсутствует. Суправитальная окраска крезиловым или метиленовым синим вызывает образование агрегатов указанных органелл, которые выявляются в виде базофильной сеточки (лат. - reticulum) в цитоплазме (что обусловило название этих форм). За время созревания ретикулоцита в крови (24-48 ч) в нем завершается сборка подмембранного комплекса элементов цитоскелета, исчезает способность к эндоцитозу, утрачиваются некоторые мембранные рецепторы и возрастает содержание гемоглобина.
Содержание ретикулоцитов в крови составляет в норме у взрослого 0.7-
1% общего числа циркулирующих эритроцитов, что приблизительно соответствует уровню их обновления в течение суток. У детей оно повышено в первые дни после рождения (до 3-5%), особенно у недоношенных (6-7%), затем несколько снижается, но в течение всего первого года жизни превышает уровень, характерный для взрослых.
Увеличение содержания ретикулоцитов (до 50% и более) может происходить вследствие их усиленного выброса костным мозгом при возникновении потребности в быстром повышении числа эритроцитов, например, после массивной кровопотери, внутрисосудистого разрушения (гемолиза) или при подъеме на высоту.
Старение эритроцитов связано с: (1) нарушением целостности подмембранного цитоскелетного комплекса, (2) изменениями в самой мембране - ее химического состава, заряда, нарушением деятельности ее ионных насосов,
(3)снижением активности ферментных систем восстановления гемоглобина и
(4)изменением его состава.
Гибель эритроцитов. Старые эритроциты, утрачивая гибкость молодых, теряют способность к прохождению через наиболее узкий участок сосудистого русла человека - щелевидные поры в эндотелии венозных синусов селезенки шириной 0.5-3 мкм. В красной пульпе селезенки они подвергаются дополнительным повреждающим воздействиям, длительно пребывая при низких значениях pH и малом содержании глюкозы. Измененные вследствие метаболических нарушений гемоглобины, связываясь с молекулами белка полосы 3, вызывают их агрегацию в кластеры. К последним на поверхности плазмолеммы присоединяются иммуноглобулины (IgG), которые обеспечивают распознавание и поглощение старых эритроцитов макрофагами.
- 170 -