Материал: В-Л-Быков-Цитогогия_и_общая_гистология
Поздние явления при некрозе включают разрыв ядерной оболочки, плазмолеммы и мембран органелл, разрушение и растворение ядра, утрату базофилии набухшей цитоплазмой. исчезновение клеточных граииц и распад клетки.
Для некроза, в отличие от апоптоза не являющегося активным процессом (см. ниже), не требуется продолжающейся синтетической активности клетки, он не сопровождается активацией пулей внутриклеточной сигнализации. Продукты распада клеток попадают в межклеточные пространства, привлекают лейкоциты и макрофаги, фагоцитирующие клеточный детрит. Фагоциты, в свою очередь, выделяют разнообразные вещества, которые обусловливают активацию и приток различных клеток вследствие хемотаксиса. Описанным образом развивается и в течение определенного времени поддерживается воспалительная реакция на продукты разрушения клеток при их некрозе.
Апоптоз - физиологическая (запрограммированная) гибель клеток.
Апоптоз (от греч. apotosis - листопад) - "смерть клетки в результате самоубийства (самоуничтожения)" - активный, генетически контролируемый процесс клеточной гибели, регулируемый внутренней программой, которая запускается внешними факторами. Развитие апоптоза индуцируется особыми генами (киллерными генами), которые обеспечивают синтез ряда веществ, обусловливающих разрушение клетки. Обнаружены также "гены-спасители", экспрессия которых противодействует развертыванию программы апоптоза (наиболее изученным ингибитором апоптоза служит ген bcl-2).
Апоптоз представляет собой энергоемкий процесс и сопровождается активацией сигнальных систем в клетке. Он обычно происходит асинхронно в отдельных клетках или мелких клеточных группах, разделенных численно превосходящими жизнеспособными клетками. Апоптоз наблюдается в различных тканях человека и животных в норме, патологии, эмбриональном развитии и у взрослого.
Сигналы, запускающие генетическую программу апоптоза, обладают специфичностью для клеток различных тканей. К наиболее общим индуцирующим сигналам относятся:
(1) Нарушение баланса регуляторных воздействий, поддерживаю пщх нормальную дифференцировку и функциональную активность клеток, например, вследствие дефицита стимулирующих факторов (гормонов, факторов роста, некоторых цитокинов), потери контакта с другими клетками или компонентами межклеточного вещества и др. К этой же категории сигналов, вероятно, относятся и те, что возникают в клетке при ее естественном старении;
- 96 -
(1)Воздействие физиологических активаторов (индукторов) апоптоза -
ФНО (фактора некроза опухолей), ИФНу (интерферона-у), ГФРР (трансформирующего фактора роста-β), молекулы Fas, глюкокортикоидов, некоторых интерлейкинов. Этот эффект тканеспецифичен: некоторые из указанных веществ в клетках одних тканей индуцируют апоптоз, тогда как в других являются его ингибиторами (факторами выживания);
(2)Воздействие разнообразных повреждающих физических и хими-ческих факторов (гипертермии, гипоксии, оксидантов, токсинов, ишемии, облучения) умеренные по интенсивности, которые при большей интенсивности приводят к развитою некроза. Причиной апоптоза обычно служат вызывемые действием указанных факторов неустранимые повреждения ДНК (см. выше) или резкие метаболические сдвиги;
(3)Некоторые инфекции, в особенности, вирусные.
Структурно-функциональные изменения клеток при апоптозе. На наиболее ранних этапах развития апоптоза до возникновения структурных изменений в клетках в их цитоплазме в течение латентного периода длительностью до 12 ч происходит синтез ферментов, которые необходимы для осуществления гибели /снетки. Эту стадию, однако, проходит большее число клеток, нежели погибает в конечном итоге, так как часть из них выживает благодаря "спасению" в результате активации особых "генов-спасителей" и действия специфических трофических факторов.
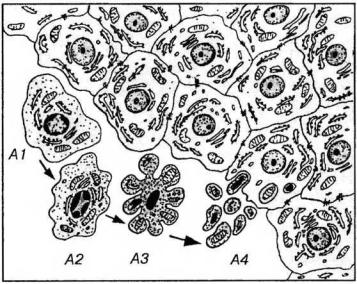
Наиболее ранним морфологическим проявлением апоптоза, выявляемым на электронно-микроскопическом уровне, служит утрата клетками специализированных структур на их поверхности (например, микроворсинок и межклеточных соединений), их отделение от соседних (рис. 3-24). Развитие апоптоза морфологически на светооптическом уровне также проявляется уплотнением ядра (в котором накапливаются крупные глыбки хроматина), конденсацией цитоплазмы, которая уплотняется, сморщивается и уменьшается в размерах (отчего аноптоз был назван также "сморщивающим некрозом" - shrinkage necrosis в англоязычной литературе). Уплотнение цитоплазмы приводит ко все более компактному расположению органелл, которые при апоитозе, в отличие от некроза, сохраняют свою целостность.
Изменения в ядре при апоптозе обусловлены активацией эндо-генной Са2 + - зависим ой эндонуклеазы, что приводит к упорядоченному расщеплению геномной ДНК в межнуклеосомных участках на отдельные нуклеосомные сегменты. Хроматин укладывается в ядре в виде крупных
полулуний, после чего ядро распадается на фрагменты,
- 97 -
окруженные мембраной. Таким образом, в отличие от некроза, изменения ядра при апоптозе включают только кариопикноз и своеобразный кариорексис (без разрушения кариолеммы); кариолизис отсутствует.
Рис. 3-24. Морфологические изменения клеток при апоптозе. А1-А4 - клетки на последовательных стадиях апоптоза: А1 - начало апоптоза - утрата клеткой соединений с соседними интактными клетками и ее отделение от них; А2 - сжатие и уплотнение цитоплазмы и ядра, изменение формы клетки, распределение гетерохроматина в виде полулуний под кариолеммой; АЗ - нарастающее сжатие и уплотнение клетки, образование вздутий и выростов на ее поверхности, кариопикноз; А4 - распад клетки на фрагменты, окруженные плазмолеммой (апоптозные тела) и их фагоцитоз соседними интактными клетками.
При прогрессировании апоптоза нарастающая конденсация цитоплазмы сочетается с изменением формы клетки - она образует многочисленные крупные вздутия и выпячивания (поверхность клетки при этом как бы "вскипает"), а также "кратеры" (по-видимому, в участках слияния с плазмолеммой пузырьков, образующихся из цистерн грЭПС).
Образование и удаление апоптозных тел. Выпячивания, содержащие жизнеспособные органеллы, а также фрагменты ядра, отшнуровываются, формируя крупные окруженные мембраной фрагменты округлой или овальной формы - апоптозные тела. Образование апоптозных тел связано с преобразованиями цитоскелета: в частности, перешнуров-
- 98 -
ка цитоплазмы происходит с участием пучков актиновых микрофиламентов, разрушение которых блокирует ход апоптоза. Число и размеры образующихся клеточных фрагментов (апоптозных тел) варьируют в широких пределах и обычно тем значительнее, чем крупнее разрушающаяся клетка. В некоторых случаях клетка сморщивается целиком, превращаясь в одно сферическое апоптозное тело. Апоптозные тела быстро захватываются соседними метками посредством фагоцитоза и перевариваются ими. Некоторые тела могут разрушаться внеклеточно, другие же поглощаются местными фагоцитами. Нейтрофилы в фагоцитозе апоптозных тел не участвуют, воспалительная реакция отсутствует.
Процесс апоптоза развивается сравнительно быстро и обычно длится от нескольких минут до нескольких часов (в среднем, морфологически регистрируемые его стадии - от начала конденсации хроматина до полного переваривания апоптозных тел, занимают 1-3 ч).
Биохимические процессы при апоптозе. Сигнал, запускающий апоптоз,
инициирует внутриклеточную каталитическую реакцию, которая включает ряд этапов:
(1)передачу сигнала в клеточное ядро (обеспечивается различными механизмами, опосредованными ионами Са2+, фосфолипазой, тирозинкиназой, протеинкиназами А и С, цАМФ, сфингомиелином/перамидом и др. молекулами),
(2)активацию "летальных", или "киллерных" генов, ответственных за развертывание программы апоптоза,
(3)включение процессов транскрипции и трансляции (в результате которых осуществляется синтез апоптоз-специфических белков),
(4)активацию ряда ферментных систем, вызывающих необрати-мые изменения в ядре и цитоплазме клетки.
Деятельность указанных ферментных систем на заключительном этапе биохимических преобразований при апоптозе генетически контролируется и координируется, разворачиваясь в определенном порядке. К наиболее важным ферментным системам относят (в порядке их активации):
(1)цистеиновые протеазы семейства ICE (сокращенное название от англ.
IL-1 Converting Enzyme - фермент, конвертирующий ИЛ-1), представляющие собой протеолитическую каскадную аутокаталитическую систему, а также ICEподобные протеазы;
(2)гранзимы (сериновые протеазы);
(3)эндонуклеазы, обусловливающие фрагментацию ДНК в участках между нуклеосомами с формированием цепей ДНК стандартной длины.
-99 -
Эта реакция, развивающаяся еще до протеолиза пистонов и других ядерных белков, настолько специфична, что ее используют в качестве маркера процесса апоптоза.
Значение апоптоза в развитии тканей и механизмах тканевого гомеостаза у человека
Апоптоз - один из фундаментальных и универсальных биологических механизмов тканевого гомеостаза, поэтому он в той или иной степени связан со всеми проявлениями жизнедеятельности тканей в норме и патологии. Особенно значима роль апоптоза в следующих процессах: (1) эмбриональном развитии; (2) удалении стареющих клеток в зрелых тканях; (3) инволюции зрелых тканей; (4) иммунных реакциях; (5) реакциях тканей на действие повреждающих факторов; (6) развитии ряда дегенеративных и инфекционных заболеваний; (7) опухолевом росте.
(1)апоптоз в эмбриональном развитии. Внутриутробное развитие сопровождается избыточным образованием огромного количества клеток, которые своевременно уничтожаются путем апоптоза. Наиболее активно этот процесс происходит в нервной системе, где механизмом апоптоза гибнет до 4085% нейронов различных участков ЦНС. Важнейшие процессы гисто- и органогенеза тесно связаны с индукцией апоптоза, развивающегося в соответствии с генетически определенной пространственной и временной программой. Апоптоз охватывает клетки в четко определенных участках формирующихся зачатков и органов на конкретных этапах их развития.
Проявлениями апоптоза в ходе развития являются регрессия частей эмбриональных зачатков и закладок органов, изменения их формы, процессы образования просвета в трубчатых органах, инволюция провизорных органов, разрыв плодных оболочек и др. Одним из механизмов действия тератогенов (от греч. teras - урод и genes - происшедший) - веществ, обусловливающих развитие уродств, - как предполагают, служит изменение расположения зон, в которых в норме происходит гибель клеток механизмом апоптоза (нарушение программы апоптоза);
(2)апоптоз стареющих клеток в зрелых тканях. Длительность жизни клеток в различных тканях варьирует в очень широких пределах - от нескольких часов (для лейкоцитов) или нескольких суток (для клеток кишечного эпителия) до многих лет (для кардиомиоцитов и нейронов). Она определена генетически и связана с характером популяции, к которым принадлежат эти клетки (см. выше). Старение клеток, неза-
- 100 -