Материал: Морфология бактерий. Спиртовое брожение
Морфология бактерий. Спиртовое брожение
Оглавление
Введение
1. Морфология бактерий
2. Спиртовое брожение
Список использованной
литературы
Введение
Микробиология ― наука о мельчайших, невидимых невооруженным глазом организмах, названных микробами или микроорганизмами. Она изучает закономерности их жизни и развития, а также изменения, вызываемые ими в организме людей, животных, растений и в неживой природе. Развитие микробиологии, как и других научных дисциплин, находится в тесной зависимости от способов производства, запросов практики, общего прогресса науки и техники.
Целью микробиологии как науки есть изучение систематики, морфологии (формы и строения) и физиологии (жизнедеятельности) микроорганизмов, методов их выделения и распознавания, а также выяснения их значения в природе и возможностей применения в различных сферах деятельности человека.
Микробиологический контроль
на пищевых производствах представляет собой все методы исследования и контроля,
связанные с определением степени бактериальной обсеменённости контролируемого
объекта, а также методы количественного учёта микрофлоры.
1.
Морфология бактерий
Форма бактерий
Подавляющее большинство известных бактерий имеют форму или сферы (шаровидные), или цилиндра (палочковидные), или спирали. Шаровидные бактерии (рис. 1.) бывают одиночными (кокки), соединенными по две клетки (диплококки), по четыре клетки (тетракокки), в длинные цепочки (стрептококки), в пакеты (сарцины), в виде скоплений неправильной формы (стафилококки). Палочковидные бактерии (рис. 1.) различаются по величине отношения длины клетки к ее поперечному размеру. У коротких палочек это отношение так мало, что их трудно отличить от кокков они подразделяются на бактерии (не образующие споры) и бациллы (образующие споры). Бактерии спиралевидной формы характеризуются разным числом витков спириллы имеют от одного до нескольких витков, вибрионы выглядят как изогнутые палочки, их можно рассматривать как неполный виток спирали.
С развитием микроскопической техники и усовершенствованием методов подготовки препаратов открыты другие экзотические формы бактерий. Некоторые бактерии имеют вид сомкнутого или разомкнутого кольца, у некоторых видны клеточные выросты (простеки), число которых колеблется от 1 до 8 и больше, обнаружены бактерии червеобразной формы, похожие на кристаллы, и т.п.
Строение бактериальной клетки
Бактериальная клетка имеет очень сложную гетерогенную и вместе с тем строго упорядоченную структуру. В общих чертах строение бактериальной клетки не отличается от строения клетки высших организмов. Клетка как универсальная единица жизни оказалась настолько совершенной формой организации живой материи, что в процессе эволюции от одноклеточных до высших многоклеточных организмов она сохранила все основные черты своего строения, а, следовательно, и функции.
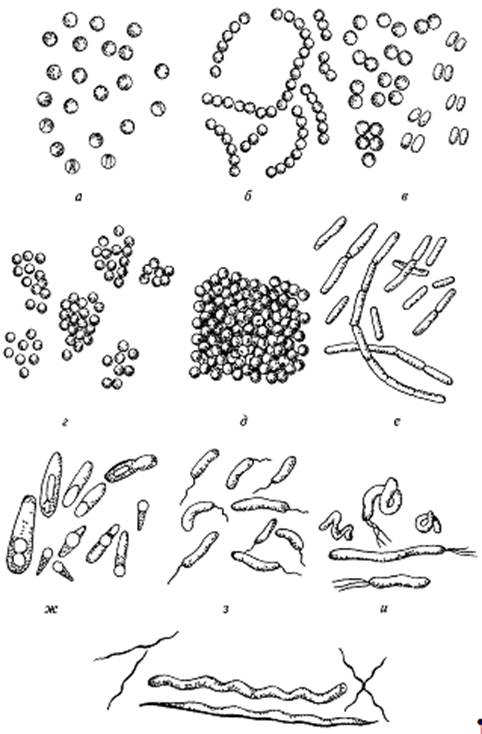
Рисунок 1. Формы бактерий. Шаровидные: а - микрококки, б - стрептококки,
в - диплококки, г - стафилококки, д - сарцины; палочковые: е - бактерии, ж -
быциллы, з,и - извитые, к - спириллы.
На рис. 2. представлена схема строения бактериальной клетки известного цитолога В.И. Бирюзовой. Форма бактериальной клетки определяется жесткой (ригидной) клеточной стенкой, которая придает клетке определенную, наследственно закрепленную внешнюю форму. На клеточной стенке бактерий находятся так называемые поверхностные структуры: капсула, жгутики, половые ворсинки, реснички. Под клеточной стенкой расположена цитоплазматическая мембрана (ЦПМ), которая отграничивает цитоплазму клетки. Цитоплазматическая мембрана вместе с цитоплазмой называется протопластом. Все слои, располагающиеся с внешней стороны от цитоплазматической мембраны, называют клеточной оболочкой.
Клеточная стенка.
У прокариот клеточная стенка состоит из пептидогликана, которого нет в
эукариотных клетках. В зависимости от строения клеточной стенки прокариоты
подразделяются на две группы: грамположительные и грамотринательные. Такое
подразделение основано на различии в способе окраски, предложенном в 1884 г.
датским ученым X. Грамом. Клеточные стенки грамположительных и
грамотрицательных прокариот резко различаются как по химическому составу, так и
по ультраструктуре.
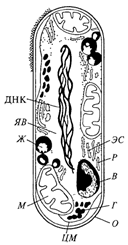
Рис. 2. Схема строения бактериальной клетки: О - оболочка клетки; ЦМ -
цитоплазматическая мембрана; М - митохондрия (мезосома); Ж- жировые
включения; ЯВ - ядерная вакуоль; ДНК - нити ДНК; ЭС - эргастоплазматическая
система; Р - рибосомы; В - волютин; Г - гликоген
В клеточной стенке грамположительных бактерий содержится 50... 90 % пептидогликана, грамотрицательных бактерий - 1... 10 % пептидогликана. Кроме пептидогликана в клеточной стенке грамположительных прокариот содержатся уникальные химические соединения - тейхоевые кислоты. На долю клеточной стенки прокариот приходится от 5 до 50 % сухих веществ клетки.
Клеточная стенка прокариот выполняет разнообразные функции: механически защищает клетку от воздействий окружающей среды, обеспечивает поддержание ее внешней формы, дает возможность клетке существовать в гипотонических растворах. В клеточной стенке расположены каналы, или диффузионные поры, для пассивного транспортирования веществ и ионов в клетку.
Клеточная стенка препятствует проникновению в клетку токсических веществ. На внешней стороне клеточной стенки расположено много макромолекул, контактирующих с окружающей средой: специфические рецепторы для фагов, антигены, макромолекулы, обеспечивающие межклеточные взаимодействия при конъюгации, а также между патогенными бактериями и клетками и тканями высших организмов.
Поверхностные структуры.
У бактерий снаружи клеточной стенки есть капсула (рис. 3.) - слизистое
образование, обволакивающее клетку, сохраняющее связь с клеточной стенкой и
имеющее аморфное строение. В зависимости от толщины капсулы бывают микрокапсулы
(толщина меньше 0,2 мкм) и макрокапсулы (толщина больше 0,2 мкм). Капсулы
защищают клетку от механических повреждений, высыхания, создают дополнительный
осмотический барьер, служат препятствием для проникновения фагов. Иногда
капсула служит источником запасных питательных веществ. Слизь помогает
прикреплению клеток к различным поверхностям. В настоящее время способность
некоторых бактерий синтезировать капсулы (своеобразные внеклеточные полимеры)
используют на практике в качестве заменителей плазмы крови и для получения
синтетических пленок.

Рис 3. Капсулы у бактерий
Многие бактерии неподвижны, если же они способны передвигаться, то это
движение обеспечивается жгутиками - структурами, расположенными на поверхности
клеток. Число, размеры и расположение жгутиков, как правило, являются
признаком, постоянным для Данного вида (рис. 4.), и имеют таксономическое
значение. Без жгутиков способны передвигаться только скользящие бактерии и
спирохеты. Обычно толщина жгутика составляет 15-20 нм, длина 3-15 мкм. Бактерии
со жгутиками могут двигаться очень быстро, например Bac. megaterium со скоростью 16 мм/мин, Vibro cholerae -
12 мм/мин.
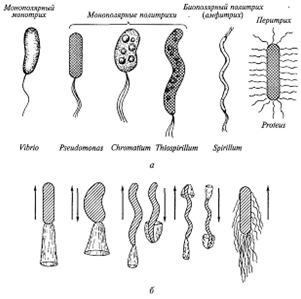
Рис. 4. Основные типы расположения жгутиков (а) и типы движения бактерий (б)
При полярном расположении жгутиков они действуют подобно корабельному винту и проталкивают клетку в окружающей жидкой среде. Вращательное движение жгутика происходит за счет базального тела. Жгутики вращаются сравнительно быстро. У спирилл они совершают около 3000 об/мин, что близко к скорости среднего электромотора. Вращение жгутиков вызывает и вращение клетки с 1/3 этой скорости в противоположном направлении.
Перитрихиально расположенные жгутики Е. coli работают как один хорошо скоординированный спиральный пучок и проталкивают клетку через среду обитания (рис. 4.).
Изучение жгутиков в электронном микроскопе показало, что они состоят из
трех частей (рис. 5.). Основную массу жгутика составляет длинная спиральная
нить (фибрилла), переходящая у поверхности клеточной стенки в утолщенную
изогнутую структуру - крючок. Нить с помощью крючка прикреплена к базальному
телу, которое представляет собой систему из двух или четырех колец (L, Р, Sw.
M), нанизанных на стержень, являющийся продолжением крючка.
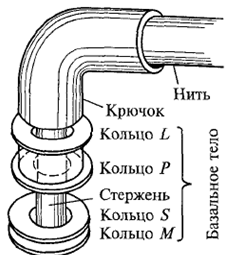
Рис. 5. Структура жгутика
В последнее время достигнуты большие успехи в расшифровке механизма
движения прокариот. Прокариотная клетка обладает механизмом, позволяющим
превращать электрохимическую энергию непосредственно в механическую. Кроме
жгутиков на клеточной стенке прокариотной клетки могут быть половые ворсинки и
реснички в виде различной длины выростов (рис. 6).
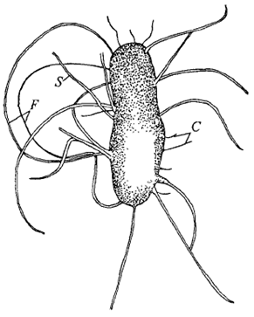
Рис. 6. Типы волосинок E.coli: F - жгутики, S -
половые ворсинки (F-like sexpili), C -
реснички.
Цитоплазматическая мембрана.
Под клеточной стенкой расположена цитоплазматическая мембрана, являющаяся обязательным структурным элементом любой клетки, нарушение целостности которого приводит к потере клеткой жизнеспособности. На долю ЦПМ приходится 8... 15 % сухого вещества клетки. ЦПМ - это белково-липидный комплекс и небольшое количество углеводов.
ЦПМ выполняет разнообразные функции с помощью специальных переносчиков, называемых транслоказами. Через мембрану осуществляется специальный перенос различных органических и неорганических молекул и ионов.
В ЦПМ локализованы многие ферменты. ЦПМ является основным барьером, обеспечивающим избирательное поступление в клетку и выход из нее разнообразных веществ и ионов.
У прокариот описаны локальные впячивания ЦПМ, которые называются мезосомами. Мезосомы различаются размерами, формой и локализацией в клетке. Считается, что с мезосомами связано усиление энергетического метаболизма клеток.
Цитоплазма.
Содержимое клетки, окруженное ЦПМ, называется цитоплазмой. Цитоплазма имеет гомогенную консистенцию и содержит набор растворимых РНК, ферментов, продуктов и субстратов метаболических реакций. В цитоплазме расположены разнообразные структуры: рибосомы, генетический аппарат (ДНК) и включения разной химической природы и функционального назначения.
Рибосомы - рибонуклеопротеидные частицы размером 15-20 нм. Их число в клетке зависит от интенсивности процесса синтеза белка. В быстрорастущей клетке Escherichia coli содержится приблизительно 15000 рибосом. Синтез белка осуществляется агрегатами, состоящими из рибосом, молекул информационных и транспортных РНК, называемых полирибосомами. Генетический аппарат прокариотной клетки представлен одной молекулой ДНК, имеющей форму ковалентно замкнутого кольца и получившей название бактериальной хромосомы. Длина молекулы ДНК в развернутом виде может составлять более 1 мм, т.е. почти в 1000 раз превышать длину бактериальной клетки. Генетический аппарат прокариотной клетки называют нуклеоидом.
В цитоплазме прокариот расположены различные включения, часть которых выполняет функцию запасных питательных веществ, представленных полисахаридами, липидами, полипептидами, полифосфатами, отложениями серы. Полисахариды - это гликоген, крахмал, гранулеза (крахмалоподобное вещество). Чаще встречаются у представителей анаэробных споровых бактерий группы клостридиев. В неблагоприятных условиях они используются в качестве источников углерода и энергии. Липиды накапливаются в виде гранул, состоящих из полимера β-оксимасляной кислоты. У некоторых прокариот, окисляющих углеводороды, поли-β-оксимасляная кислота составляет до 70 % сухого вещества клетки.
Липиды служат для клетки хорошим источником углерода и энергии. Полифосфаты, также накапливающиеся в виде гранул, называются волютиновыми и используются клетками как источник фосфора. Для бактерий, осуществляющих хемосинтез за счет окисления сероводорода, характерно накопление в клетках молекулярной серы. Все накопленные (запасные) вещества, представленные в виде высокомолекулярных полимерных молекул, отграничены от цитоплазмы белковой мембраной.
Пигменты бактерий
Колонии многих бактерий ярко окрашены. Способность к синтезу пигментов обусловлена генетически. Среди пигментов бактерий встречаются каротиноиды, феназиновые красители, пирролы, азахиноны, антоцианы и др.
Пигменты защищают клетки от светового повреждения и используют свет для фотосинтеза. У многих микроорганизмов образование пигмента идет только на свету. Например, ярко-красная окраска колоний у Serratia marcescens обусловлена присутствием пигмента продигиозина. Бактерии Pseudomonas indigofem, Cotynebacterium insidiosum, Arthrobacter atrocyaneus и др. синтезируют индигоидин - нерастворимый в воде синий пигмент, выделяемый в среду. Chromobacterium violaceum образует сине-фиолетовый пигмент виолацеин, нерастворимый в воде. Виолацеин является производным индола, образующегося при окислении триптофана. Pseudomonas aeruginosa образует пигмент никс-пиоцианин. Различные штаммы псевдомонад образуют такие пигменты, как Феназин-1-карбоновую кислоту, оксихлорофин, иодинин, а иногда все пигменты вместе.
Все пигменты относятся к вторичным метаболитам, т.е. они не принадлежат к тем соединениям, которые имеются у всех организмов, и являются производными обычных метаболитов или структурных компонентов клетки. Некоторые пигменты обладают антибиотическими свойствами, так что многие пигментированные микроорганизмы являются продуцентами антибиотиков.
Рост и способы размножения бактерий
Рост прокариотной клетки - это согласованное увеличение количества всех химических компонентов, из которых она построена. Рост является результатом множества скоординированных биосинтетических процессов, находящихся под строгим регулярным контролем, и приводит к увеличению массы и размеров клетки. Рост клетки не беспределен. После достижения определенных (критических) размеров клетка подвергается делению. Для большинства прокариот характерно равновеликое бинарное поперечное деление, приводящее к образованию двух одинаковых дочерних клеток.
У большинства грамположительных бактерий деление происходит путем синтеза поперечной перегородки, идущей от периферии к центру. Поперечная перегородка формируется из ЦПМ и пептидогликанового слоя. Наружные слои синтезируются позднее.
Клетки большинства грамотрицательных бактерий делятся путем перетяжки. Например, у Е. coli на месте деления обнаруживается постепенно увеличивающееся и направленное внутрь искривление ЦПМ и клеточной стенки.
Вариантом бинарного деления является почкование, при котором на одном из полюсов материнской клетки образуется маленький вырост (почка), увеличивающийся в процессе роста. Постепенно почка достигает размеров материнской клетки и отделяется от нее. Почкующиеся клетки подвергаются старению. При равновеликом бинарном делении материнская клетка дает начало двум дочерним клеткам, а сама исчезает. При почковании материнская клетка дает начало дочерней клетке и между ними можно обнаружить морфологические различия. Деление прокариотной клетки начинается, как правило, спустя некоторое время после завершения цикла деления ДНК.
Спорообразование бактерий
Вегетативные клетки многих прокариот для перенесения неблагоприятных условий образуют специальные клетки (эндоспоры), обладающие повышенной устойчивостью. В основе морфологического дифференцирования лежат биохимические процессы, запрограммированные соответствующей генетической информацией. Образование эндоспор происходит у прокариот и грибов.
Эндоспора формируется внутри материнской клетки (спорангия), обладает специфическими структурами: многослойными белковыми покровами, наружной и внутренней мембранами, кортексом (рис. 7.). Эндоспоры устойчивы к повышенным температурам, дозам радиации, которые детальны для вегетативных клеток. К спорообразующим бактериям относится большое число прокариот из 15 родов, среди которых есть палочковидные, сферические, спириллы и нитчатые организмы. Все они имеют клеточную стенку, характерную для грамположительных прокариот. В каждой бактериальной клетке образуется, как правило, одна эндоспора.