Материал: микра от 104 группы
36. Силосование кормов. Микробиологические процессы при разных способах силосования. Методы регулирования процесса силосования.
Силосование, или заквашивание, — способ консервирования зеленого корма, при котором растительную массу хранят во влажном состоянии в ямах, траншеях или специальных сооружениях — силосных башнях.
Корм, более или менее спрессованный и изолированный от доступа воздуха, подвергается брожению, приобретает кислый вкус, становится мягче, несколько изменяет цвет (бурая окраска), но остается сочным.
Способы силосования кормов
Известны два способа силосования: холодный и горячий.
При холодном способе силосования созревание силоса идет при умеренном повышении температуры, доходящем в некоторых слоях корма до 40 °С; оптимальной температурой считается 25—30 °С. При таком силосовании скошенную растительную массу, если нужно, измельчают, укладывают до отказа в кормовместилище, утрамбовывают, сверху как можно плотнее укрывают для изоляции от воздуха.
Холодный способ силосования наиболее распространен, что объясняется как сравнительной его простотой, так и хорошим качеством получающегося корма.
При горячем способе силосное сооружение заполняют по частям. Зеленую массу на один-два дня рыхло укладывают слоем около 1—1,5 м. При большом количестве воздуха в ней активизируются микробиологические и ферментные процессы, в результате чего температура корма поднимается до 45—50 °С. Затем укладывают второй слой такой же толщины, как и первый, и он, в свою очередь, подвергается разогреванию.
Так слой за слоем заполняют все силосохранилище. Самый верхний слой корма утрамбовывают и плотно прикрывают для защиты от воздуха. В связи с тем, что силосохранилище при горячем способе силосования обычно делают небольших размеров, на верхний слой силосуемого корма помещают груз.
Разогревание растительной массы связано с потерей иногда значительной части питательных веществ корма. В частности, резко уменьшается переваримость белков. Поэтому горячее силосование не может считаться рациональным способом сохранения растительной массы. Общие потери сухих веществ корма при холодном силосовании не должны превышать 10—15%, во втором достигают 30% и более
Горячий способ силосования допустим лишь для квашений грубостебельных, малоценных кормов, которые после разогревания лучше поедаются скотом.
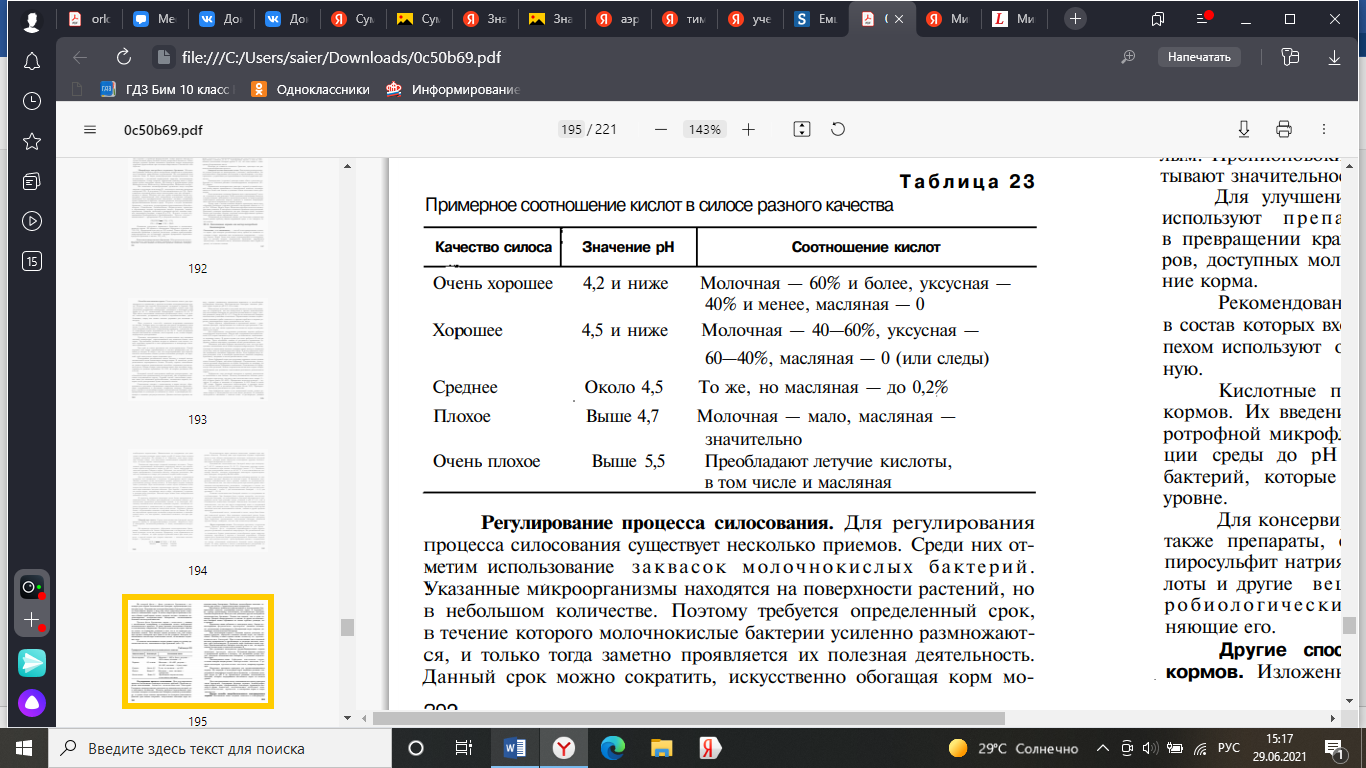
Для того чтобы процесс силосования проходил интенсивно, необходимо создать благоприятные условия для развития молочнокислых бактерий, быстро повышающих кислотность корма до уровня (рН 4—4,2), при котором невозможно развитие нежелательных процессов брожения. В результате жизнедеятельности этих бактерий сахар силосуемых растений в большей своей части превращается в молочную кислоту.
Герметизация и кислотность силоса — главнейшие факторы, определяющие его стойкость при хранении. Снижение по тем или иным причинам кислотности корма неминуемо ведет к его порче.
Микрофлора силоса
Среди молочнокислых бактерий силоса имеются кокки и неспорообразующие палочки: Streptococcus lactis, S. thermophilus, Lactobacillus plantarum, L. brevis, являющиеся анаэробами.
6С5Н10О5 = 8С3Н6О3 + ЗС2Н4О2
Пентоза= Молочная кис + Уксусная кислота кис
В силосе могут развиваться кислотоустойчивые дрожжи, не оказывающие вредного влияния на качество корма. В правильно заложенной заквашиваемой массе дрожжи активно не размножаются. Это объясняется тем, что они не могут расти при низком уровне окислительно-восстановительного потенциала, создаваемого в силосе молочнокислыми бактериями.
Развитие маслянокислых бактерий связано со следующими их особенностями. Эти бактерии более строгие анаэробы, чем молочнокислые бактерии, но неустойчивы к высокой кислотности и прекращают расти при реакции среды, близкой к рН 4,7—5,0, как и большинство гнилостных бактерий. Накопление масляной кислоты нежелательно, так как она имеет неприятный запах и содержащий ее корм скот поедает плохо. При подобном брожении корма кроме масляной кислоты накапливаются амины, аммиак и другие вредные продукты.
В растительной массе, заложенной в силос, могут быть бактерии кишечной группы. Они вызывают гнилостный распад белка, а сахар превращают в малоценные для консервирования продукты. При нормально протекающем силосовании бактерии кишечной группы быстро отмирают, так как они некислотоустойчивы.
Фазы созревания силоса
Первая фаза созревания заквашиваемого корма характеризуется развитием смешанной микрофлоры. На растительной массе начинается бурное размножение разнообразных групп микроорганизмов, внесенных с кормом в силосное помещение. Обычно первая фаза брожения бывает кратковременной. Окончание первой, или предварительной, фазы брожения связано с подкислением среды, угнетающей деятельность большей части микрофлоры корма. К этому времени в силосе устанавливаются анаэробные условия, так как весь кислород уже израсходован аэробами.
Во второй фазе — фазе главного брожения — основную роль играют молочнокислые бактерии, продолжающие подкислять корм. Большинство неспорообразующих бактерий погибает, но бациллярные формы в виде спор могут длительное время сохраняться в заквашенном корме. В начале второй фазы брожения в силосе обычно преобладают кокки, которые позднее сменяются палочковидными молочнокислыми бактериями, отличающимися большой кислотоустойчивостью.
Третья фаза брожения корма — конечная — связана с постепенным отмиранием в созревающем силосе возбудителей молочнокислого процесса. К этому времени силосование подходит к естественному завершению. Быстрота подкисления корма зависит не только от количества углеводов в нем, но и от структуры растительных тканей. Чем быстрее отдают растения сок, тем скорее идет процесс квашения при одних и тех же условиях. Быстроте заквашивания способствует измельчение массы, облегчающее отделение сока.
Регулирование процесса силосования
Для регулирования процесса силосования существует несколько приемов. Среди них отметим использование заквасок молочнокислых бактерий. Указанные микроорганизмы находятся на поверхности растений, но в небольшом количестве. Поэтому требуется определенный срок, в течение которого молочнокислые бактерии усиленно размножаются, и только тогда заметно проявляется их полезная деятельность. Данный срок можно сократить, искусственно обогащая корм молочнокислыми бактериями. Особенно целесообразно внесение заквасок при работе с трудносилосуемым материалом.
Предложена технология приготовления и использования бактериальных заквасок, улучшающих качество корма. В большинстве случаев рекомендуют использовать молочнокислую бактерию Lactobacillus plantarum. Иногда к ней добавляют другого возбудителя молочнокислого брожения. Готовят как жидкие, так и сухие закваски. Данный микроорганизм в отличие от других молочнокислых бактерий может сбраживать не только простые углеводы, но и крахмал.
Предлагают также добавлять в силосуемую массу, бедную моносахаридами, ферментные препараты (мальтаза, целлюлаза), разлагающие полисахариды и обогащающие корм сахарами, доступными молочнокислым бактериям.
При силосовании кормов с большим запасом углеводов, например кукурузы, образуется слишком кислый корм, что нежелательно. Поэтому в таких случаях готовят закваску из пропионовокислых бактерий. При ее использовании часть молочной кислоты превращается в пропионовую и уксусную кислоты, которые слабо диссоциируют. В результате корм становится менее кислым. Пропионовокислые бактерии полезны еще и тем, что вырабатывают значительное количество витамина В12.
Для улучшения силосуемости труднозаквашиваемых кормов используют препарат милазы. Указанный фермент участвует в превращении крахмала в мальтозу, что увеличивает резерв сахаров, доступных молочнокислым бактериям, и усиливает подкисление корма.
Рекомендованы также буферные кислотные смеси, в состав которых входят разные минеральные кислоты. С успехом используют органические кислоты, например, муравьиную.
Кислотные препараты пригодны для труднозаквашиваемых кормов. Их введение в силосуемый корм подавляет развитие сапротрофной микрофлоры первой фазы брожения, а снижение реакции среды до рН 4 не препятствует развитию молочнокислых бактерий, которые поддерживают кислотность корма на низком уровне.
Для консервирования плохо заквашиваемых кормов пригодны также препараты, содержащие формиат кальция, метабисульфит, пиросульфит натрия, сульфаминовую, бензойную, муравьиную кислоты и другие вещества, подавляющие побочные микробиологические процессы в силосуемом корме и сохраняющие его.
37. Круговорот углерода и роль в нём микроорганизмов. Значение аэробных и анаэробных процессов превращения соединений углерода в природе и для сельского хозяйства.
Круговорот углерода обеспечивается созидательным процессом - фотосинтезом зеленых растений, с одной стороны, а с другой стороны, процессами распада безазотистых веществ до конечных продуктов, обусловленными жизнедеятельностью микроорганизмов. Зеленые растения образуют за год органическое вещество, содержащее 1,5·1014 т углерода и соответственно 1,4·1020 кал энергии. Содержание углекислоты в атмосфере поддерживается в некоторой степени также извержениями вулканов, дыханием животных и растений и сжиганием топлива, например каменного угля, сохранившего огромные запасы солнечной энергии до наших дней.
Некоторое количество углекислоты усваивается гетеротрофными, а также хемосинтезирующими микробами. Последние используют образующуюся при окислении неорганических веществ (аммиак, сера) энергию для восстановления углекислоты. Но этот процесс по сравнению с фотосинтезом очень невелик.
Микроорганизмы, минерализуя органические вещества, вновь возвращают углерод в атмосферу в форме углекислоты и этим делают возможным дальнейшее развитие жизни на Земле.
Поскольку растительные и животные остатки содержат белки, углеводы и жиры, то разложение их в природных условиях протекает одновременно, но осуществляется большей частью разными физиологическими группами микроорганизмов.
Разложение органических безазотистых веществ осуществляется микроорганизмами двумя путями: или посредством брожения, или путем окисления.
При кислородном фотосинтезе аэробные цианобактерии и зелёные растения основную часть окисленной формы углерода (СО2) переводят в восстановленное состояние, характерное для органических соединений (например, глюкозу), а восстановленную форму кислорода (Н20) окисляют до 02. Хотя анаэробные пурпурные и зелёные бактерии могут восстанавливать С02 до органических веществ, окисляя отличные от воды соединения (NH3, NO2, Н2, Fe2+, восстановленные соединения серы), вклад этих процессов в общую фиксацию СО, незначителен. В результате фотосинтетической фиксации С02 образуются сахара и другие соединения. Основная масса фиксированного углерода растений откладывается в виде полимерных углеводов (крахмал, целлюлоза). Поэтому сахара играют ведущую роль в питании всех живых организмов, нуждающихся в органической пище (организмы-гетеротрофы), и служат предпочтительными питательными веществами для большинства гетеротрофных микроорганизмов.
В анаэробных условиях органические соединения расщепляются путём брожения (дрожжи, молочнокислые бактерии, пропионовокислые бактерии, бактерии семейства Entembacteriaceae), либо окисляются в процессе анаэробного дыхания при наличии акцепторов водорода. В роли акцепторов водорода выступают нитраты, сульфаты, карбонаты, фумараты, Fe3+: соответственно выделяют денитрифицирующие, сульфатредупирующие, метанобразующие бактериями. Метанобразующие бактерии (Methanobacterium, Melhanococcus, Methanosarcina) — строгие анаэробы, составляющие последнее звено анаэробной пищевой цепи. Выделяемый ими метан в аэробных условиях может окислятся до С02 метилотрофными бактериями (Methylomonas, Methytosinus, Melhylococcus).
В процессе белкового обмена в организме животных и человека накапливается мочевина, которая выводится во внешнюю среду с мочой. Уробактерии сбраживают мочевину с образованием аммиака, углекислоты и воды. Таким образом, органический азот мочевины, которая выделяется животными и людьми в громадных количествах (более 50 млн т мочевины в год), становится доступным для растениями.
Аммиачные соли, образующиеся при распаде белков или расщеплении мочевины, окисляются в азотнокислые соли, которые усваиваются растениями. Этот процесс именуется н и т р и ф и к а ц и е й. Она протекает в две фазы: 1) окисление аммиачных солей до солей азотистой кислоты, осуществляемое нитрозобактериями, которые были открыты русским учёным С. Н. Виноградским; 2) окисление солей азотистой кислоты (нитритов) в соли азотной кислоты (нитраты), происходящее под влиянием нитробактерий.
Процессы, обратные нитрификации, - д е н и т р и ф и к а ц и я, осуществляются микроорганизмами, восстанавливающими соли азотной кислоты в соли азотистой кислоты. В результате этого образуется азот, который улетучивается в атмосферу. Почва при этом обедняется, и плодородие её снижается. Частым перепахиванием и рыхлением создают условия усиленной аэрации почвы, чем уменьшают или прекращают денитрификацию. Процессы денитрификации компенсируются деятельностью азотусваивающих или азотфиксирующих бактерий, которые усваивают атмосферный азот и переводят его в соединения, доступные для питания растений. К ним относят некоторые свободноживущие почвенные бактерии, азотобактер, а также клубеньковые бактерии, которые живут на корнях бобовых растений, находясь с последними в симбиотических отношениях.
Для удобрения почвы применяют бактериальные препараты: нитрагин – чистую культуру клубеньковых бактерий в стерилизованной почве и азотобактерин – культуру азотобактера на нейтральном торфе.