Материал: fiza_3_semestr_2
8. Альвеолярная вентиляция легких.
Газовая смесь, поступившая в легкие при вдохе, распределяется на две части. Одна из них не принимает участие в газообмене, т.к. заполняет воздухоносные пути (анатомически мертвое пространство). Другая часть (альвеолярный объем) поступает в респираторный отдел (альвеолярные протоки, мешочки и альвеолы), где принимает участие в газообмене. Она обеспечивает вентиляцию альвеолярного пространства.
Особенности альвеолярной вентиляции:
-интенсивность обновления газового состава, определяемая соотношением альвеолярного объема и альвеолярной вентиляции.
-изменения альвеолярного объема (увеличение/уменьшение размера вентилируемых альвеол, либо изменение кол-ва альвеол, участвующих в вентиляции).
-различия внутрилегочных характеристик сопротивления и эластичности, приводящее к асинхронности альвеолярной вентиляции.
-поток газов в альвеолу или из нее определяется механическими характеристиками легких и дыхательных путей, а также силами (или давлением), воздействующими на них. Механические характеристики обусловлены сопротивлением дыхательных путей потоку воздуха и эластическими св-ва легочной паренхимы.
Размеры альвеолярного пространства таковы, что смешивание газов в альвеолярной единице происходит практически мгновенно как следствие дыхательных движений, кровотока и диффузии.
Неравномерность альвеолярной вентиляции обусловлена и гравитационным фактором-разницей транспульмонального давления в верхних и нижних отделах грудной клетки. В вертикальном положении в нижних отделах это давление выше примерно на 8 см.вод.ст. Апико-базальный градиент всегда присутствует независимо от степени наполнения легких→определяет наполнение воздухом альвеол в разных отделах легких.
В норме вдыхаемый газ смешивается мгновенно с альвеолярным газом. Состав газа в альвеолах практически гомогенен в любую респираторную фазу и в любой момент вентиляции.
Любое повышение альвеолярного транспорта кислорода и углекислого газа (напр. при физических нагрузках) сопровождается повышение градиентов концентрации газов, которые способствуют возрастанию их смешивания в альвеолах. Нагрузка стимулирует альвеолярное смешивание путем повышения потока вдыхаемого воздуха и возрастания кровотока, повышает альвеолярно-капиллярный градиент давления для кислорода и углекислого газа.
9. Диффузия газов.
Газообмен — совокупность процессов, обеспечивающих переход кислорода внешней среды в ткани живого организма, а углекислого газа из тканей во внешнюю среду.
Перемещение газов осуществляется под влиянием разности парциальных давлений и напряжений этих газов в каждой из сред организма.
Парциальное давление кислорода в воздухе, заполняющем альвеолы легких, около 100 мм рт. ст., а его напряжение в венозной крови, притекающей к легким, около 40 мм рт. ст. Вследствие разности давлений кислород из альвеол направляется в кровь, где связывается с гемоглобином эритроцитов. Парциальное давление углекислого газа в альвеолярном воздухе составляет 40 мм рт. ст., а его напряжение в притекающей к легким венозной крови — 48 мм рт. ст. Вследствие разности давлений углекислый газ переходит в альвеолы.
В артериальной крови, притекающей к тканям, напряжение кислорода выше, чем в тканях, а напряжение углекислого газа, наоборот, значительно ниже. Вследствие этого кислород переходит из крови в ткани и включается в цикл метаболических процессов, а углекислый газ, в избытке содержащийся в тканях, переходит в кровь и переносится затем в лёгкие. Процесс газообмена происходит непрерывно до тех пор, пока существует разность парциальных давлений и напряжений газов в каждой из сред, участвующих в газообмене решающим фактором, обусловливающим непрерывность газообмена, является постоянство газового состава альвеолярного воздуха.
Величина газообмена является показателем интенсивности окислительных процессов, протекающих в тканях. Об уровне газообмена можно судить и по величине минутной вентиляции легких. При спокойном дыхании через легкие проходит около 8000 мл воздуха в 1 мин. При физических и эмоциональных напряжениях, различных заболеваниях, сопровождающихся усилением окислительных процессов в тканях, легочная вентиляция возрастает.
Вентиляционно-перфузионные отношения в разных отделах легкого.
Кровоток в капиллярах легких и легочная вентиляция неодинаковы в различных отделах и зависят от положения тела.
Основное влияние на распределение перфузии в легких оказывает гравитация, что обусловлено низким АД в системе малого круга кровообращения (15-20 мм рт. ст.). Поэтому при любом положении тела в пространстве нижние отделы легких по сравнению с верхними будут иметь больший кровоток.
Зависимость перфузии от сил гравитации более выражена, чем у вентиляции, что определяет и характер изменения вентиляционно-перфузионных отношений по направлению от верхушек к основанию легких. Нормальная альвеолярная вентиляция (VA) у взрослых составляет ~ 4 л/мин, а общая легочная перфузия (Q) ~ 5 л/мин. Следовательно, отношение величин вентиляции и перфузии будет равно 4/5, или 0,8. Изменение отношения YA /Q будет отражать степень гипервентиляции (гипоперфузии) или гиперперфузии (гиповентиляции) в целом легком или в его отдельных зонах.
Распределение вентиляции зависит от нескольких факторов. Основным является растяжимость легочной ткани, которая неодинакова в различных легочных зонах.
10. Транспорт кислорода.
Транспорт О2 осуществляется в физически растворенном и химически связанном виде. Физические процессы, т. е. растворение газа, не могут обеспечить запросы организма в О2.
Согласно закону Фика, газообмен О2 между альвеолярным воздухом и кровью происходит благодаря наличию концентрационного градиента О2 между этими средами. В альвеолах легких парциальное давление О2 составляет 13,3 кПа, или 100 мм рт.ст., а в притекающей к легким венозной крови парциальное напряжение О2 составляет примерно 5,3 кПа, или 40 мм рт.ст. Давление газов в воде или в тканях организма обозначают термином «напряжение газов» и обозначают символами Ро2, Рсo2.
Транспорт О2 начинается в капиллярах легких после его химического связывания с гемоглобином. Гемоглобин (Нb) способен избирательно связывать О2 и образовывать оксигемоглобин (НbО2) в зоне высокой концентрации О2 в легких и освобождать молекулярный О2 в области пониженного содержания О2 в тканях. При этом свойства гемоглобина не изменяются и он может выполнять свою функцию на протяжении длительного времени.
Гемоглобин переносит О2 от легких к тканям. Эта функция зависит от двух свойств гемоглобина: 1) способности изменяться от восстановленной формы, которая называется дезоксигемоглобином, до окисленной (Нb + О2 à НbО2) с высокой скоростью (полупериод 0,01 с и менее) при нормальном Рог в альвеолярном воздухе; 2) способности отдавать О2 в тканях (НbО2 à Нb + О2) в зависимости от метаболических потребностей клеток организма.
Зависимость степени оксигенации гемоглобина от парциального давления Ог в альвеолярном воздухе графически представляется в виде кривой диссоциации оксигемоглобина, или сатурационной кривой. Плато кривой диссоциации характерно для насыщенной О2 (сатурированной) артериальной крови, а крутая нисходящая часть кривой — венозной, или десатурированной, крови в тканях.
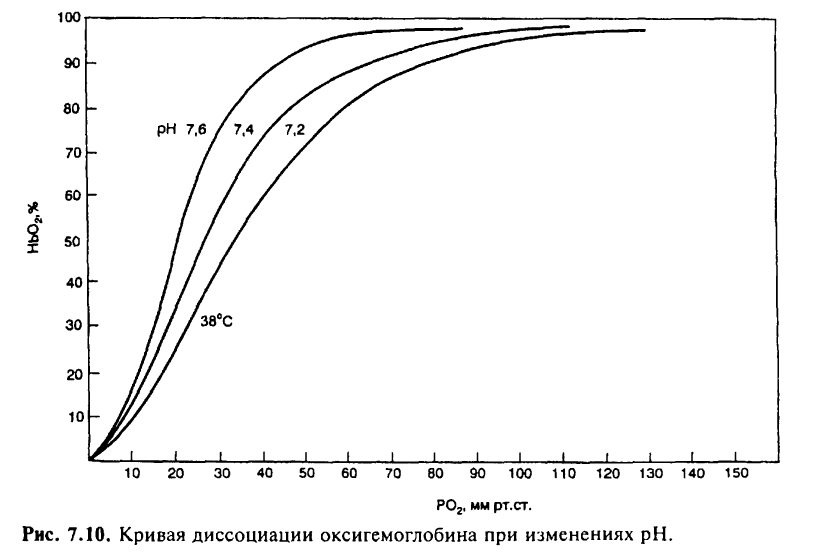
На сродство кислорода к гемоглобину влияют различные метаболические факторы, что выражается в виде смещения кривой диссоциации влево или вправо. Сродство гемоглобина к кислороду регулируется важнейшими факторами метаболизма тканей: Ро2 pH, температурой и внутриклеточной концентрацией 2,3-дифосфоглицерата. Величина рН и содержание СО2 в любой части организма закономерно изменяют сродство гемоглобина к О2: уменьшение рН крови вызывает сдвиг кривой диссоциации соответственно вправо (уменьшается сродство гемоглобина к О2), а увеличение рН крови — сдвиг кривой диссоциации влево (повышается сродство гемоглобина к О2). Например, рН в эритроцитах на 0,2 единицы ниже, чем в плазме крови. В тканях вследствие повышенного содержания СО2 рН также меньше, чем в плазме крови. Влияние рН на кривую диссоциации оксигемоглобина называется «эффектом Бора».
 Рост
температуры уменьшает сродство
гемоглобина к О2. В работающих мышцах
увеличение температуры способствует
освобождению О2. Уменьшение температуры
тканей или содержания 2,3-дифосфоглицерата
вызывает сдвиг влево кривой диссоциации
оксигемоглобина.
Рост
температуры уменьшает сродство
гемоглобина к О2. В работающих мышцах
увеличение температуры способствует
освобождению О2. Уменьшение температуры
тканей или содержания 2,3-дифосфоглицерата
вызывает сдвиг влево кривой диссоциации
оксигемоглобина.
Метаболические факторы являются основными регуляторами связывания О2 с гемоглобином в капиллярах легких, когда уровень O2, рН и СО2 в крови повышает сродство гемоглобина к О2 по ходу легочных капилляров. В условиях тканей организма эти же факторы метаболизма понижают сродство гемоглобина к О2 и способствуют переходу оксигемоглобина в его восстановленную форму — дезоксигемоглобин. В результате О2 по концентрационному градиенту поступает из крови тканевых капилляров в ткани организма.
Оксид углерода (II) — СО, способен соединяться с атомом железа гемоглобина, изменяя его свойства и реакцию с О2. Очень высокое сродство СО к Нb (в 200 раз выше, чем у О2) блокируют один или более атомов железа в молекуле гема, изменяя сродство Нb к О2.
Под кислородной емкостью крови понимают количество Ог, которое связывается кровью до полного насыщения гемоглобина. При содержании гемоглобина в крови 8,7 ммоль*л-1 кислородная емкость крови составляет 0,19 мл О2 в 1 мл крови (температура 0oC и барометрическое давление 760 мм рт.ст., или 101,3 кПа). Величину кислородной емкости крови определяет количество гемоглобина, 1 г которого связывает 1,36—1,34 мл О2. Кровь человека содержит около 700—800 г гемоглобина и может связать таким образом почти 1 л О2. Физически растворенного в 1 мл плазмы крови О2 очень мало (около 0,003 мл), что не может обеспечить кислородный запрос тканей. Растворимость О2 в плазме крови равна 0,225 мл*л-1*кПа-1.
Обмен О2 между кровью капилляров и клетками тканей также осуществляется путем диффузии. Концентрационный градиент О2 между артериальной кровью (100 мм рт.ст., или 13,3 кПа) и тканями (около 40 мм рт.ст., или 5,3 кПа) равен в среднем 60 мм рт.ст. (8,0 кПа). Изменение градиента может быть обусловлено как содержанием О2 в артериальной крови, так и коэффициентом утилизации О2, который составляет в среднем для организма 30— 40%. Коэффициентом утилизации кислорода называется количество О2, отданного при прохождении крови через тканевые капилляры, отнесенное к кислородной емкости крови.
С другой стороны, известно, что при напряжении О2 в артериальной крови капилляров, равном 100 мм рт.ст. (13,3 кПа), на мембранах клеток, находящихся между капиллярами, эта величина не превышает 20 мм рт.ст. (2,7 кПа), а в митохондриях равна в среднем 0,5 мм рт.ст. (0,06 кПа).
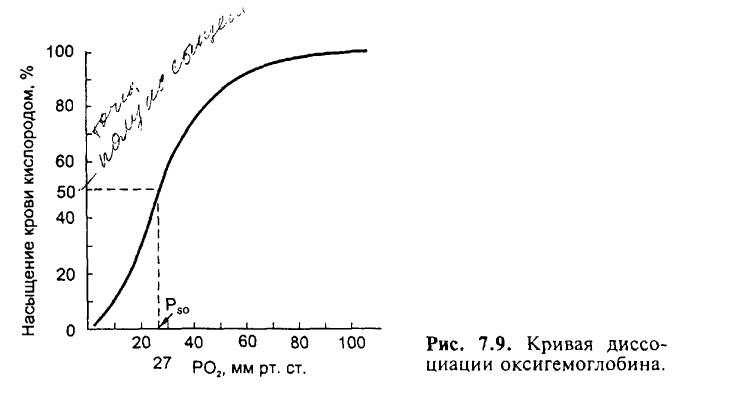
11. Кривая диссоциации оксигемоглобина.
Реакция взаимодействия кислорода с гемоглобином подчиняется закону действующих масс. Это означает, что соотношение между количествами гемоглобина и оксигемоглобина зависит от содержания физически растворенного О2 в крови; последнее же, согласно закону Генри-Дальтона, пропорционально напряжению О2. Процент оксигемоглобина от общего содержания гемоглобина называют кислородным насыщением (SО2) гемоглобина. Если гемоглобин полностью дезоксигенирован, то SО2 = 0%; если же весь гемоглобин превратился в оксигемоглобин, то SО2 = 100%. В соответствии с законом действующих масс насыщение гемоглобина кислородом зависит от напряжения О2. Графически эту зависимость отражает так называемая кривая диссоциации оксигемоглобина. Эта кривая имеет S-образную форму. Расположение кривой диссоциации оксигемоглобина зависит от ряда факторов (см. ниже). Наиболее простым показателем, характеризующим расположение этой кривой, служит так называемое напряжение полунасыщения (50%), т.е. такое напряжение О2, при котором насыщение гемоглобина кислородом составляет 50%. В норме (при рН=7,4 и t=37°C) полунасыщение артериальной крови составляет около 26 мм рт.ст. (3,46 кПа).
Причины S-образной формы кривой диссоциации оксигемоглобина до конца не ясны. Если бы каждая молекула гемоглобина присоединяла только одну молекулу О2, то кинетика этой реакции графически описывалась бы гиперболой. Именно такая гиперболическая кривая диссоциации характерна, например, для реакции соединения кислорода с красным мышечным пигментом миоглобином, аналогичной реакции оксигенации гемоглобина. Строение миоглобина сходно со структурой одной из четырех субъединиц гемоглобина, поэтому молекулярные массы этих двух веществ соотносятся как 1:4. Поскольку в состав миоглобина входит лишь одна пигментная группа, одна молекула миоглобина может присоединить только одну молекулу О2. Исходя из вполне правдоподобного предположения о том, что S-образная форма кривой диссоциации НbО2 обусловлена связыванием одной молекулой гемоглобина четырех молекул О2, Эдер выдвинул так называемую гипотезу промежуточных соединений. Согласно этой гипотезе, присоединение четырех молекул О2 к гемоглобину происходит в несколько стадий, причем каждая из этих стадий влияет на равновесие следующей реакции. Таким образом, реакция соединения кислорода с гемоглобином описывается четырьмя константами равновесия, что и объясняет сигмоидную форму кривой диссоциации оксигемоглобина.
В то же время возможно и другое объяснение, согласно которому существуют две формы гемоглобина-оксигенированная и дезоксигенированная, переходящие одна в другую в результате конформационных перестроек. Если предположить, что параметры равновесия реакций оксигенации для этих двух форм гемоглобина различны, то с позиции данной гипотезы можно объяснить S-образную форму кривой диссоциации НbО2
Биологический смысл формы кривой диссоциации оксигемоглобина. Конфигурация кривой диссоциации оксигемоглобина имеет важное значение с точки зрения переноса кислорода кровью. В процессе поглощения кислорода в легких напряжение О2 в крови приближается к таковому в альвеолах. У молодых людей РО2 артериальной крови составляет около 95 мм рт.ст. (12,6 кПа). При таком напряжении насыщение гемоглобина кислородом составляет примерно 97%. С возрастом (и в еще большей степени при заболеваниях легких) напряжение О2 в артериальной крови может значительно снижаться, однако, поскольку кривая диссоциации оксигемоглобина в правой ее части почти горизонтальна, насыщение крови кислородом уменьшается ненамного. Так, даже при падении РО2 в артериальной крови до 60 мм рт.ст. (8,0 кПа) насыщение гемоглобина кислородом равно 90%. Таким образом, благодаря тому, что области высоких напряжений кислорода соответствует горизонтальный участок кривой диссоциации оксигемоглобина, предупреждается cyщественное снижение насыщения артериальной крови кислородом.
Крутой наклон среднего участка кривой диссоциации оксигемоглобина свидетельствует об очень благоприятных условиях для отдачи кислорода тканям. При изменении локальной потребности в кислороде он должен высвобождаться в достаточном количестве в отсутствие значительных сдвигов РО2 в артериальной крови. В состоянии покоя РО2 в области венозного конца капилляра равно приблизительно 40 мм рт.ст. (5,3 кПа), что соответствует примерно 73% насыщения. Если в результате увеличения потребления кислорода его напряжение в венозной крови падает лишь на 5 мм рт.ст. (0,7 кПа), то насыщение гемоглобина кислородом снижается не менее чем на 7%; высвобождающийся при этом О2 может быть сразу же использован для процессов метаболизма.