Материал: 9. Мембрана Документ Microsoft Office Word
![]()
.
|
Дифузія лінійно залежить від |
|
|
|
концентрації С0. Це випадок прос- |
|
|
|
тої дифузії. Процес полегшеної |
Рис. 3.18. Залежність початкової |
|
|
дифузії характеризується тим, що |
|
|
|
швидкості дифузії речовини j |
|
|
|
він: |
|
|
|
від концентрації С0 за полегшеної дифузії |
|
– специфічний для певних молекул;
– швидкість його вища за швидкість простої дифузії;
– досягає насичення;
– чутливий до дії певних інгібіторів.
Провідність мембрани визначається концентрацією наявних в ній
141
носіїв струму та їх рухливістю. Низька діелектрична проникність ліпідів (ε = 2…3) дуже несприятлива для впровадження заряджених частинок у мембрану.
Можна показати [29], що величина енергетичного бар’єра в мем-брані зменшується, а отже, проникність мембрани для іона зростає зі збільшенням радіуса іона r і наближенням величини діелектричної про-никності мембрани εм до діелектричної проникності води εв. На цих фі-зичних принципах і ґрунтується перенесення іонів іонофорами. Іонофо-ри можуть утворювати з іоном комплекс великого розміру – переносники, або формувати пори в мембрані, які заповнені водою, – канали.
Перший механізм полягає в тому, що молекула переносника захоп-лює речовину (тим самим немовби збільшуючи його радіус), що транс-портується на одному боці мембрани і разом з ним рухається через лі-підний бішар на другий бік мембрани. Це – перенесення за допомогою рухомого переносника. Прикладом рухомих переносників є іонофорні антибіотики, що здійснюють таким чином транспорт іонів лужних ме-талів, наприклад, валіноміцин транспортує іони калію.
Другим механізмом полегшеної дифузії є транспорт за допомогою спеціальних пор, яке називають також естафетним перенесенням.
-
цьому випадку пора може бути утворена декількома послідовно роз-
міщеними молекулами, між якими передаються іони. Наприклад, анти-біотик граміцидин А переносить іони Na+.
Полегшеною дифузією пояснюється перенесення через біологічні мембрани амінокислот, цукру та інших біологічно значущих речовин.
Відмінності полегшеної дифузії від простої:
– речовина за участю переносника переноситься значно швидше;
– полегшена дифузія має властивість насичення;
– за меншої дифузії спостерігається конкуренція перенесених речо-вин у тих випадках, коли переносником переносяться різні речовини; так, із цукру глюкоза переноситься швидше ніж фруктоза, фруктоза швидше ніж ксілоза, а ксілоза швидше ніж арабіноза;
142
– є речовини, які блокують полегшену дифузію і утворюють міцний комплекс з молекулами переносника.
Фільтрацією називають рух розчину через пори в мембрані під дією градієнта тиску. Швидкість перенесення у процесі фільтрації під-
коряється закону Пуазейля:
|
d |
|
p p |
, |
|
|
|
|
1 |
2 |
|
||
|
|
|
|
|
||
|
dt |
|
|
w |
|
|
![]()
де dυ/dt – об’ємна швидкість перенесення розчину; w – гідравлічний опір. Явище фільтрації відіграє важливу роль у процесах перенесення
води через стінки кровоносних судин.
Осмос – проста дифузія води місця з її більшою концентрацією в місця з меншою концентрацією води. Осмос відіграє велику роль у ба-гатьох біологічних явищах. Явище осмосу зумовлює гемоліз еритроци-тів у гіпотонічних розчинах.
3.5.2. Поняття активного транспорту речовини
Активний транспорт – це перенесення речовини з місць з мен-шим значенням електрохімічного потенціалу в місця з його більшим значенням. Активний транспорт у мембрані супроводжується зрос-танням енергії Гібса, він не може здійснюватися мимовільно, а лише за рахунок витрати енергії, яка запасена в макроергічних зв’язках АТФ.
Активний транспорт речовин через біологічні мембрани має вели-чезне значення. За рахунок активного транспорту в організмі створю-ються градієнти концентрацій, градієнти електричних потенціалів, гра-дієнти тиску тощо, які підтримують життєві процеси, тобто з погляду термодинаміки активне перенесення утримує організм у нерівноважно-му стані, підтримує життя.
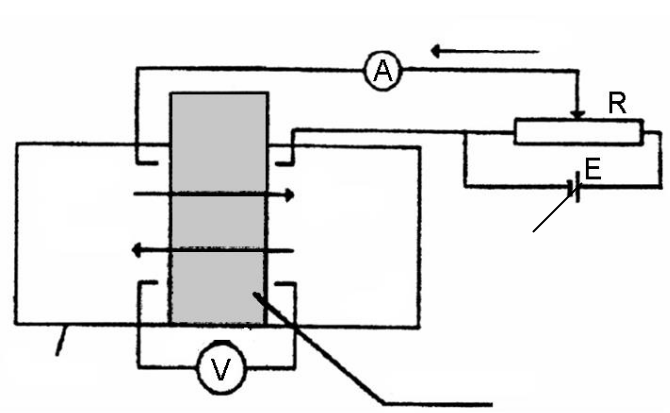
Активний транспорт через біологічні мембрани досліджував дан-ський учений Г. Уссінг (1949 р.) у дослідах з перенесення іонів натрію через шкіру жаби (рис. 3.19) [56]. Перенесення ґрунтувалося на вимірю-
143
ванні активного транспорту електричним методом (методом короткого замикання). Якщо розділити дві камери з однаковими розчинами Рінге-ра шкірою жаби, то між її зов-
|
|
|
І ≠ 0 |
нішньою (мукозною) та внут- |
|
||||||
|
|
|
|
|
|||||||
|
|
|
1 |
рішньою (серозною) поверх- |
|
||||||
|
|
|
|
|
|||||||
|
Na22 |
|
jm.вн |
нями |
виникає |
різниця |
|
||||
|
jm.зов |
|
2 |
потенціалів, яка приблизно до- |
|
||||||
|
|
|
|
|
|
||||||
|
|
Na24 |
рівнює 100 мВ (потенціал на |
|
|||||||
|
|
|
|
||||||||
|
Камера |
Δφ=0 |
Шкіра жаби |
зовнішній |
поверхні |
буде |
|
||||
|
|
від’ємним). При цьому спосте- |
|
||||||||
|
з електролітом |
|
|||||||||
|
|
|
|||||||||
|
Рис. 3.19. Схема експериментальної |
рігалися потоки іонів |
натрію |
|
|||||||
|
|
установки Уссінга: |
крізь шкіру жаби: від зовніш- |
|
|||||||
|
|
А – амперметр, V – вольтметр; |
|
||||||||
|
|
ньої до внутрішньої поверхні і |
|
||||||||
|
1– реостат; 2– конденсат; 3– гальванічний |
|
|||||||||
|
|
|
елемент |
від внутрішньої до зовнішньої |
|
||||||
поверхні. Відношення потоків іонів у разі пасивного транспорту вира-жається рівнянням Уссінга:
![]()
|
j |
|
C |
e |
|
|
ZF |
|
|
|
k |
e |
RT |
. |
|
||||
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
||
|
j |
|
|
i |
|
|
|
|
|
|
|
C |
|
|
|
|
|
||
|
|
|
|
k |
|
|
|
|
|
![]()
![]()
Регулюючи струм у компенсувальному колі, можна досягти різниці потенціалів до нуля. Тоді в системі має настати рівновага, оскільки не-має градієнтів концентрацій іонів і потенціалу між камерами, розділе-ними шкірою жаби. В таких умовах пасивне перенесення іонів згідно з критерієм Уссінга неможливе, сумарний потік дорівнює нулю.
Проте під час проведення експериментів було виявлено [94], що струм, який тече в компенсаційному колі, протікає і крізь шкіру. От-же, відбувається одностороннє перенесення іонів. Методом мічених атомів було показано, що потік іонів натрію від зовнішньої до внут-рішньої поверхні більший від потоку в протилежному напрямку. Та-ким чином, струм усередині епітелію створюється активним перене-сенням іонів натрію.
144
Згідно із сучасним уявленням активно транспортуються іонні насоси біологічних мембран – спеціальні білкові комплекси (транспортні АТФ). Відомі три типи електрогенних іонних насосів. Комплекс К+-Na+-АТФ за рахунок енергії гідролізу однієї молекули АТФ у клітину переносяться два іони калію і виносяться з клітини три іони натрію. Отже, насос генерує електричний струм через мембрану, тому К+–Na+-насос – електрогенний. Са+–АТФ переносить два іони кальцію, а Н+-насос – два протони з вико-ристанням енергії однієї молекули АТФ.
-
більшості моделей активного транспорту передбачається процес сполучення хімічної реакції гідролізу АТФ (скалярного процесу) з на-прямленим просторовим переміщенням (векторним процесом). У цьому випадку білок або білковий комплекс повинен виконувати як фермента-тивну, так і транспортну функції. Отже, у простому випадку він може містити один активний центр і один центр скріплення іонів. Для забез-печення транспорту іонів між двома мембранами передбачається, що центр скріплення іонів може поперемінно контактувати з обома розчи-нами, розділеними мембраною. Це, наприклад, мінорні рухи, які відкри-вають та закривають канали, сполучені з розділеними мембраною роз-чинами, або повернення всієї молекули ферменту в площину мембрани, що зумовлюють пересування центра скріплення іонів через мембрану. Можливі також зміни просторової форми макромолекули для перемі-
щення центра скріплення йонів.
Як показали дослідження, білок К+-Na+-АТФ існує в двох різних конформаціях – Е1 та Е2, при цьому Е1 має високу спорідненість з Na+ і низьким – з К+ (місця скріплення іонів орієнтовані до цитоплазми). Ця конформація білка має високу спорідненість з молекулами АТФ. Кон-формація Е2 ферменту має високу спорідненість з К+ і низьку – з Na+ (місця скріплення обернені назовні). При цьому Na+ потрібен для швид-кого фосфоритування, а К+ – для швидкого дефосфоритування.
Перехід іонів К+ із зв’язаного з ферментом стану у вільний стан ві-дбувається на завершальному п’ятому етапі. Цей процес індукується
145
скріпленням АТФ з алостеричним центром ферменту, який має низьку спорідненість.
Таким чином, у процесі активного транспорту молекула АТФ вико-нує дві функції – субстрати та алостеричного ефектора.
Запропоновані моделі будови, згідно з якими ці білкові комплекси містять 6, 8 або 10 трансмембранних α-спіральних сегментів. Є велика гідрофільна ділянка, що містить домени, де зв’язуються нуклеотиди і де відбувається фосфоритування. При цьому зміна конформації, що супро-воджується гідролізом АТФ, зумовлює зміну спорідненості відповідних центрів білка з іонами.
Створені за допомогою систем активного транспорту градієнти концентрацій речовин утворюють на мембрані різницю хімічного або електрохімічного потенціалу, за рахунок чого можуть переноситися ін-ші речовини проти градієнтів їх концентрацій. Такий транспорт, джере-лом енергії якого є не безпосередньо АТФ або енергія окиснювально-відновних реакцій, а градієнт концентрацій інших речовин називають вторинно-активним, або зв’язаним.
-
три види вторинно-активного транспорту іонів: уніпорт, симпорт
-
антипорт.
-
разі уніпорту за рахунок існування на мембрані градієнта елект-ричного потенціалу здійснюється однонапрямлений транспорт зарядже-них частинок у бік меншого значення потенціалу. У процесі антипорту (обмінного транспорту) здійснюється транспорт однаково заряджених іонів двох типів у різних напрямках. Згідно з механізмом симпорту (ко-транспорту) здійснюється транспорт протилежно заряджених іонів в одному напрямі. При цьому один з іонів транспортується за градієнтом концентрацій, а другий – за градієнтом електричного потенціалу, який створюється транспортом першого іона.
Одним з найбільш вивчених випадків вторинно-активного транс-порту незаряджених молекул є всмоктування глюкози в стінки кишеч-нику. Тут транспорт глюкози здійснюється проти градієнта концент-
146
рації, але не за рахунок енергії АТФ, а завдяки градієнту концентрації іншої речовини, у цьому випадку – іонів натрію. Подібні системи тра-нспорту існують і для багатьох інших речовин, наприклад вуглеводів і амінокислот, що дуже важливо, оскільки для них немає специфічних насосів.