Материал: 9. Мембрана Документ Microsoft Office Word
. Транспорт речовини через біологічні мембрани
Процеси, що відбуваються в клітині, такі, як збудження, обмін речо-вин, синтез АТФ, підтримання постійного іонного складу, вміст води та інші, залежать від перенесення речовин через біологічні мембрани. Зміна швидкості перенесення речовин призводить до порушення водного сольо-вого обміну, збудливості клітин, біоенергетичних та інших процесів.
Процеси вибірного перенесення речовин через біомембрани нази-вають транспортом. Розрізняють активний та пасивний транспорт. Тра-нспорт речовин через клітинну мембрану називають пасивним, якщо на
137
нього не витрачається клітинна хімічна енергія АТФ, і активним, якщо витрачається енергія АТФ.
3.5.1. Поняття пасивного транспорту речовини
Пасивний транспорт – це перенесення речовини з місць з великим зна-ченням електрохімічного потенціалу до місць з його меншим значенням.
Пасивний транспорт витрачається зі зменшенням енергії Гібса, і тому цей процес може перебігати самостійно без витрати енергії.
Густина потоку речовини jm під час пасивного транспорту підкоря-ється рівнянню Теорелла:
jm UC ddx ,

![]()
де U – рухливість частинок; С – їх концентрація. Знак «–» показує, що перенесення відбувається у бік спадання густини потоку.
Густина потоку речовини – це величина, яка чисельно дорівнює кіль-кості речовини масою m, яка переноситься за одиницю часу t через одини-цю площі S поверхні, перпендикулярної до напрямку перенесення:
|
j |
|
m |
. |
|
|
|
|
|||
|
m |
|
St |
|
|
|
|
|
|
|
![]()
Підставивши останній вираз у рівняння Теорелла, отримують для розбавлених розчинів, якщо μ0 = const, рівняння Нернста–Планка:
|
j |
URT |
dC |
UCZF |
d |
. |
|
|
|
|
|
||||
|
m |
|
dx |
|
dx |
|
|
|
|
|
|
|
|
![]()
![]()
Отже, можуть бути дві причини перенесення речовини за пасивним транспортом: градієнт концентрації і градієнт електричного потенціалу. Знаки «–» перед градієнтами показують, що градієнт концентрації зу-мовлює перенесення речовини від місць з більшою концентрацією до місць з його меншою концентрацією, а градієнт електричного потенціа-лу зумовлює перенесення позитивних зарядів від місць з великим потен-ціалом до місць з меншим потенціалом.
138
-
разі відсутності електролітів (Z = 0) або електричного поля (d/dx = 0) рівняння Теорелла переходить у рівняння:
|
j |
URT |
dC |
. |
|
|
|
|
|||
|
m |
|
dx |
|
|
|
|
|
|
|
![]()
Згідно зі співвідношенням Ейнштейна коефіцієнт дифузії
D RTu,
де R – універсальна газова стала; Т – абсолютна температура; u – рухомість речовини в розглядуваному середовищі.
-
результаті отримане рівняння, що описує просту дифузію, є зако-
ном Фіка:
|
j |
D |
dC |
, |
|
|
|
|
|||
|
m |
|
dx |
|
|
|
|
|
|
|
![]()
де D – коефіцієнт дифузії, см2.с; С – концентрація, моль∙см3. Знак «–» у рівнянні вказує на те, що дифузія відбувається в напрямку, протилеж-ному вектору градієнта концентрації, тобто у напрямку менших значень концентрації.
Класифікацію основних видів пасивного транспорту через мембра-ну показано на рис. 3.17.
Основними способами пасивного проникнення речовин через біомем-брани є проста і полегшена дифузія [63]. Дифузія – мимовільне переміщен-ня речовини з місць з більшою концентрацією в місця з меншою концент-рацією речовини унаслідок хаотичного теплового руху молекул.
На практиці за умов, що градієнт концентрації С = const, корис-туються формулою, що пов’язує густину потоку речовини через мем-брану з концентраціями цієї речовини не всередині мембрани, а зовні в розчинах біля поверхонь мембрани С1 та С2:
-
DC DC2 C1 K PC2 C1, l0

де Р – коефіцієнт проникності мембрани, м/с; К – коефіцієнт розчин-ності; l0 – товщина мембрани.
139
|
|
|
|
|
|
|
|
|
Пасивний транспорт |
|
|
|
|
|
|
||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||||||||||||||||||||
|
|
|
Осмос |
|
|
|
|
|
|
|
|
|
|
|
Фільтрація |
|
|
||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
Проста |
|
|
Полегшена |
|
|
|
|||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
дифузія |
|
|
|
дифузія |
|
|
|
||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||||||||||||||||||||||||||||
|
|
Крізь |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
З рухомим |
|
|
|
|
|
З фіксованим |
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||
|
|
ліпідний |
|
|
|
|
|
переносником |
|
|
|
|
переносником |
|
|
||||||||||||||||||||||||||||||||||||||||||||||
|
|
шар |
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||||||||||||||||||||||||||
|
|
Крізь пори |
|
|
|
|
Крізь |
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||||||||||||||||||||||||
|
|
в ліпідному |
|
|
|
білкову |
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||||||||||||||||||||||||||||
|
|
шарі |
|
|
|
|
пору |
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||||||||||||||||||||
![]()

![]()
![]()
Рис. 3.17. Класифікація видів пасивного транспорту
Коефіцієнт проникності мембрани залежить від властивостей мем-
брани і переносних речовин. Коефіцієнт проникності
|
P |
DK |
. |
|
|
l |
|
||
|
|
|
|
![]()
Коефіцієнт проникності тим більший, чим більший коефіцієнт ди-фузії (менша в’язкість мембрани), чим тонша мембрана (менше l) і чим краще речовина розчиняється в мембрані (чим більший К). Високу про-никність мембрани мають неполярні речовини (органічні жирні кисло-ти, ефіри), а низьку – полярні, водорозчинні речовини (солі, цукор, амі-нокислоти, спирти).
-
разі нестаціонарної дифузії концентрація речовини в будь-якій точці вздовж осі х залежить від часу. Можна показати [32], що квадрат відстані, на яку розповсюджуються частинки дифундувальної речовини, лінійно залежить від часу:
![]() x2
x2
![]() 4Dt.
4Dt.
![]()
![]()
Коефіцієнт дифузії визначають за розмірами та формою молекул. Для малих молекул у воді D ≈ 10–5 см2/с. Дифузія сферичних частинок у рідині описується рівнянням Стокса–Ейнштейна:
|
D |
k |
|
|
|
B |
T , |
|
|
|
6r |
|
||
|
|
|
|
![]()
де kв – стала Больцмана; r – радіус частинок; – в’язкість середовища.
140
Дифузія істотно залежить від розчинності дифундованої речовини в ліпідному бішарі. У ліпідах дифундують неполярні (ліпофільні) речови-ни, тобто такі, які не мають ні заряду, ні дипольного моменту. Тому проникна здатність неполярних органічних сполук через мембрану дуже велика. Органічні сполуки, що мають полярні групи (ОН, NО2), гірше розчиняються в ліпідах, ніж неполярні сполуки, тому і проникність мембран для них істотно менша.
За низької концентрації речовини
|
j j |
C |
, |
|
|
|
0 |
|
|||
|
max |
|
|
||
|
|
C |
|
|
|
|
|
m |
|
|
|
![]()
де Сm – концентрація речовини, за якої j = jmax /2 (рис. 3.18).
Іони та полярні молекули не дуже малих розмірів проникають у
клітину за допомогою особливих білків-переносників (іонофорів). Це по-
легшена дифузія. Початкову швид-
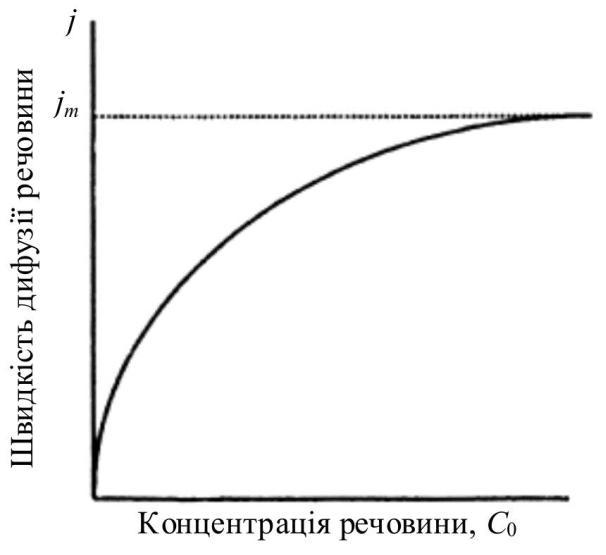
кість дифузії, коли з одного боку
мембрани концентрація речовини
-
= Сm, а з другого боку дорівнює
нулю, можна подати виразом
-
|
j |
|
C |
|
|
|
|
0 |
|
||
|
max |
|
C |
C |
|
|
|
|
|
||
|
|
|
m |
0 |
|