Курсовая работа (т): Синантропизация растений в промышленной зоне
) сообщества на своеобразных субстратах (выходы останцевых гранито-гнейсовых пород);
) экстразональные сообщества в местах контактов зональных типов растительности.
Если в ближайшие годы не будет снижено возрастающее воздействие антропогенных факторов, то бор необходимо будет отнести и к группе сообществ, сокращающих свои территории и находящихся на грани исчезновения под влиянием хозяйственной деятельности человека[14].
Учитывая, что на границах бора растительность его оказывается наиболее подверженной антропогенным изменениям, для детальных исследований были выбраны периферические участки Парижского лесничества. Почти 20 лет на землях лесничества с мая по ноябрь производится выпас овец Астафьевского совхоза Нагайбакского района. Однако на территории лесничества сохранились обширные остепненные поляны (возникшие, вероятно, на месте распространенных здесь прежде сообществ сухих остепненных сосновых лесов и редколесий в результате воздействия сплошных и выборочных рубок, пожаров и пастьбы скота) с сообществами, близкими к естественным, растительность которых принята за исходную.
На всей территории преобладают чернозем сильновыщелоченный среднегумусный маломощный среднещебенчатый и чернозем неполноразвитый среднегумусный маломощный в комплексе с выходами горных пород от 10 до 25%. Для данного района различными авторами отмечалось богатое разнообразие ассоциаций разнотравно-дерновинно-злаковых, богаторазнотравно-типчаково-ковыльных, ковыльно-разнотравных кустарниковых и каменистых степей [9, 11, 3, 4.]. Бессистемное стравливание выпасных участков свело исходное разнообразие фитоценозов в результате конвергенции растительных сообществ к сходному пастбищному травостою - однообразным и низкопродуктивным полынно-ковылково-типчаковым, луковично-мятликово-типчаковым и обыкновеннотысячелистниково-типча-ковым ассоциациям.
На участках, растительность которых принята нами за исходную, травяной покров высокорослый (80-100 см) с преобладанием злаков (Stipa capillata, реже S. pennata, S. lessingiana, Calamagrosus epigeios, Koeleria glauca, Phleum phleoides) и богатым ксеро- и мезофитным разнотравьем. Крупные дерновины злаков (диаметром 14-22 см) несколько приподняты, образуя «кочки» высотой от 4 до 10 см. Среди разнотравья много особей с генеративными побегами. Закустаренность караганой (высотой 80-120 см) и более низкорослыми (до 50 см) вишней степной, ракитником русским, шиповником обыкновенным, дроком красильным, спиреей городчатой достигает 80 %. На 100 м2 насчитывается от 35 до 44 видов, из них два-три синантропных с обилием sol. Общее проективное покрытие 95-100 %. Почва покрыта хорошо развитым войлоком из отмерших надземных органов растений[14.].
На I стадии деградации (умеренный выпас) характерен однообразный ковыльник (в основном Stipa capillata) с примесью Festuca valesiaca ssp. sulcata. Уменьшается обилие Tragapogon orientalis, Sedum stepposum, Campanula carvicaria, Aster amellus, Galium verum, Gypsophylla paniculata, Veronica longifolia, Sanguisorba officinalis. Ha 100 м2 - 29-38 видов, из них 18-26 разнотравья. Синантропных видов с обилием от sol. до sp. до шести. Общее проективное покрытие 40-60 %. Увеличение обилия злаков в составе травостоя делает растительность данных участков аспективно более однообразной по сравнению с «эталонными» участками. Почти исчезает мохово-лишайниковый покров. Степной войлок чаще отсутствует, образуется небольшой (1-2 см) слой ветоши.
На II стадии деградации (интенсивный выпас) наблюдается выпадение из травостоя Stipa pennata, S. capillata уступает позицию доминанту Festuca valesiaca ssp. sulcata, сохраняясь в обилии cop.1- sp. Характерны микрогруппировки с доминированием некоторых синантропных видов (Polygonum aviculare, Rumex acetosella, Trifolium repens, Vicia cracca, Urtica urens, Erodium cicutarium), вкрапленные в полынно (Artemisia absintium, A. sieversiana)-типчаковые, обыкновенно-тысячелистниково-типчаково-луковичномятликовые ассоциации. Уменьшается диаметр дерновин злаков до 7-11 см. На 100 м2 24-38 видов. Общее проективное покрытие 40-60 %. Появляющиеся оголенные участки почвы покрыты лишь рыхлой подстилкой из отмерших побегов данного вегетационного периода. Аспективно растительность данной стадии отличается комплексностью - зеленые пятна вейника на общем буром фоне или преобладание в аспекте пятен тысячелистника, полыни или чабреца. Местами обильно разрастается и иногда даже образует бурьянистые заросли чертополох.
При выпасе овец изменение флористического состава происходит несколько иначе, чем при выпасе крупного рогатого скота, и первая стадия дигрессии характеризуется уменьшением обилия многих видов разнотравья, а не угасанием ковылей, как это должно быть. Даже на сбоях ковыль-волосатик часто входит в число доминирующих видов. На исследованных площадках отмечено снижение жизненности ковылей и выпадение их из травостоя при большом развитии вейника, на что указывает также Г. М. Зозулин [8.].
К III стадии (чрезмерный выпас) отнесены сообщества, возникающие в местах различных типов сбоев, водопоя. Они состоят из 7-18 видов, большинство которых относится к синантропным. Распространяются однолетники: Amaranthus retroflexus, Camelina glabrata, Chenopodium album, Ch. rubrum, Euphrasia tatarica, Malva pusilla. Преобладают быстро отрастающие дерновинные злаки (Festuca valesiaca ssp. sulcata, Helictotrichon desertorum, Koeleria gracilis, Stipa capillata) и низкотравные (до 10 см) розеточные гемикриптофиты (Carex rupestris, Oxytropis spicata, Potentilla orientalis, Pulsatilla patens, P. flavescens, Veronica incana). Обглоданные кустики караганы выделяются оголенными коричневыми побегами; спирея и дрок красильный образуют после отрастания низкорослые подушки. Общее проективное покрытие снижается до 10-30 %.
Неравномерное распределение растительности, кучное и медленное передвижение стада овец, отчуждение растения на высоте 3-5 см от поверхности почвы быстро доводят многие участки пастбища до состояния крайней выбитости и появления голых пятен почвы.
Отмечен ряд общих последствий пастбищной деградации растительности:
) Происходит обеднение видового состава травянистой растительности. Число видов на 100 м2 уменьшается от 44 до 7. Упрощение и унификация растительности неизбежно сопровождаются снижением ее устойчивости по отношению ко всякого рода воздействиям. [5.]
) Происходит уменьшение численности особей и понижение жизненности кустарников (Caragana frutex, Spiraea crenata, Genista tinctoria, Cytisus ruthenicus).
) Увеличивается разреженность растительного покрова. Степень проективного покрытия уменьшается с 90 до 10%. Снижается высота травостоя до 3-5 см, преобладает низкотравье. Упрощается вертикальная структура фитоценозов, часто до двухъярусной: мятлик - типчак, чабрец - типчак.
) Упрощается эколого-ценотическая структура растительности. Увеличивается ксерофитность растительного покрова, сокращается обилие мезофильных видов (Galium verum, Thalictrum minus, Polygonatum officinale и др.). Ксерофитизации растительности способствует и изменение почвы (утаптывание, повышение капиллярности, интенсивное нагревание и потеря воды из-за оголения, вторичное засоление).
) Снижается продуктивность пастбищ. Травостой обедняется хозяйственно ценными видами, засоряется непоедаемыми, плохо поедаемыми и ядовитыми растениями (Hyoscyamus niger, Carduus crispus, Echinops sphaerocephalum, Eryngium planum, Thymus serpullum). Увеличивается численность эфемеров и однолетников, с разрастанием которых укорачивается период вегетации, что также понижает продуктивность пастбищ.
) Увеличивается число синантропных видов от 3 до 12 и обилие их от sol. до сор.2. Пастбищная деградация - одна из форм проявления синантропизации растительного покрова [5.].
Выявленные изменения растительности выпасаемых участков позволяют сделать вывод о том, что большинство пастбищных участков Парижского лесничества находится на последних стадиях деградации.
На площадках, использовавшихся под временные загоны для овец, изменяется экологическая обстановка, возникает определенный микроклимат. Почва, в отличие от растительности,, обладает способностью длительное время «нейтрализовать» неблагоприятные на нее внешние воздействия [18.]. Изучение почвенных профилей и прикопок показало, что исходные эдафические условия нивелируются образующимся напочвенным слоем. Существенно изменяются температурный и водный режимы. Темный цвет напочвенного слоя способствует интенсивному нагреванию в дневные часы, рыхлая его структура - резкому понижению температуры ночью. Увеличение перепада суточных температур вызывает интенсификацию испарения влаги из верхнего горизонта почвы. Уменьшение запасов влаги на фоне крайне неблагоприятного увлажнения, характерного для степной зоны, препятствует возобновлению растений. Накапливающиеся на овечьих стоянках экскременты - источник химического и биологического, загрязнения. Избыток удобрений вызывает засоление почвы. Моча животных увеличивает щелочность, вызывая «выгорание» растений. Развиваются эрозионные процессы, усиливающиеся при полном разрушении растительности[14.]. Они препятствуют самовосстановлению травянистых и тем более древесных сообществ. Жесткая экологическая обстановка оказывает заметное влияние на подбор видов в ходе восстановления растительности
ГЛАВА 3. ХИМИЧЕСКАЯ СТОРОНА ПРОЦЕССОВ
СИНАНТРОПИЗАЦИИ
.1 Адаптационные процессы в растительных
организмах связанные с синантропизацией
Растения, произрастающие на урбанизированных территориях, испытывают на себе постоянное влияние техногенного загрязнения. Поэтому одним из факторов, определяющих жизнеспособность зеленых насаждений в городских условиях, является их устойчивость к действию фитотоксичных соединений, загрязняющих почву и воздух.
Значительную роль в ответных реакциях растений на неблагоприятные условия среды оказывают окислительно-восстановительные процессы, в частности реакции, протекающие с участием кислородных радикалов [19.]. Образование активных форм кислорода (АФК) - синглетный кислород (О2*), супероксид-радикал (О2¯), гидроксил-радикал (ОН) и перекись водорода (Н2О2) называемое "окислительным взрывом", является одним из ранних ответов на стрессовое воздействие. Радикалы кислорода и продукты их превращения представляют серьезную угрозу для живого организма, так как могут подавлять активность ферментов, вызывать изменения нуклеиновых кислот, деградацию белков, изменять проницаемость мембран и т.д. [4].
В клетках существует динамическое равновесие между образованием АФК и их ликвидацией, которое осуществляется с помощью многокомпонентной системы антиоксидантной защиты (АОС), состоящей из низко- и высокомолекулярных компонентов. Важнейшими высокомолекулярными антиоксидантами растений, непосредственно обезвреживающими АФК, являются супероксиддисмутаза (СОД), пероксидаза (ПО), каталаза (КАТ). Ферменты-антиоксиданты, обеспечивающие комплексную защиту биополимеров от АФК, расположены в различных клеточных компартментах, имеют разную субстратную специфичность и сродство к активным формам кислорода. [7].
Кроме того, немаловажное значение в регуляции окислительных процессов отводится ферменту полифенолоксидазе (ПФО), хотя этот фермент и не входит в состав АОС [13.]. В этих работах показано, что в стрессовых условиях (при облучении, механическом повреждении, изменении химического состава окружающей среды) активность фенолоксидаз возрастает, что проявляется в образовании химических барьеров, препятствующих дальнейшему распространению АФК.
Возможно, ферменты АОС и ПФО принимают участие в регуляции метаболизма в ходе онтогенеза и имеют особое значение для растений, способствуя их быстрому приспособлению к постоянно меняющимся условиям внешней среды. Наличие нескольких ферментов, выполняющих одну и ту же каталитическую функцию, - весьма ценное свойство, расширяющее адаптационные возможности организма, что особенно важно для растений, не имеющих стабильной внутренней среды [6.].
При окислительном стрессе ферментативная АОС может становиться неэффективной. Причины этого - быстрая инактивация конститутивного пула ферментов свободными радикалами. В этих условиях повышается значение низкомолекулярных неферментативных антиоксидантов. К ним относятся различные по структуре и химическим свойствам соединения, способные взаимодействовать с кислородными и органическими радикалами и ингибировать протекание свободнорадикальных процессов в клетках. Среди них известны как гидрофобные (витамин А, каротиноиды и др.), так и гидрофильные протекторы (витамин С, фенолы, глутатион и др.). Механизм их действия состоит в том, что они "подставляют себя под удар" реактивных производных кислорода (О2*, О2¯, ОН•, Н2О2, органические радикалы и т.п.) и, окисляясь, прерывают опасную для клетки цепь реакций. Однако неферментативные низкомолекулярные антиоксиданты являются менее эффективной АОС по сравнению с ферментативной.
В ряде работ показано, что изменение активности
АОС наблюдается в ответ на действие неблагоприятных факторов среды, таких как
засуха, засоление, действие низкой температуры, повышение концентрации цинка в
среде, повышение содержание азота, загрязнение атмосферного воздуха, вирусные
инфекции и т.д
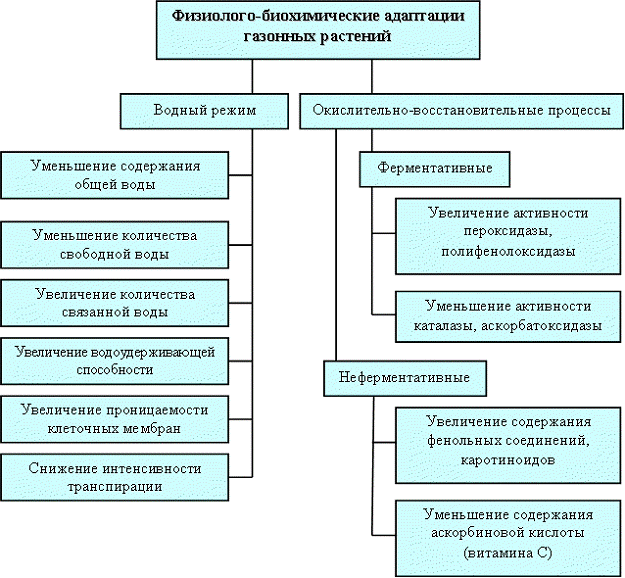
Рис.2
На основании большого экспериментального материала, полученного за последние годы, установлено, что универсальной реакцией растительной клетки на экстремальные условия внешней среды является активизация процесса перекисного окисления липидов [2], и поэтому одним из маркеров окислительного стресса растений является уровень содержания пероксидных групп. Анализ пероксидных групп в подземных органах исследованных видов выявил увеличение содержания гидропероксидов по мере загрязнения среды и возрастание количества перекисей в прегенеративном и постгенеративном периодах развития растений с его падением в генеративном периоде.
Среди ферментов АОС особая роль отводится ПО, которая способна реагировать на широкий спектр факторов, приводящих к нарушению гомеостаза АФК у растений. Под влиянием неблагоприятных воздействий среды у этого фермента меняется активность, причем повышение активности происходит параллельно с увеличением степени техногенной нагрузки на растения.
Максимальная активность ПО совпадает с периодом наиболее интенсивных метаболических процессов, происходящих во время цветения и плодоношения растений. Однако причины активации ПО загрязняющими веществами, как и многими экстремальными воздействиями абиогенной и биогенной природы, еще не достаточно ясны. Полагают, что увеличение активности этого фермента при действии стресс-факторов может быть связано с изменением соотношения его форм. Поскольку ПО в растительной клетке находится как в свободном, так и в связанном состоянии, то под влиянием стресса может происходить увеличение фонда свободных форм за счет их перехода из связанного состояния. Вместе с тем, немаловажное значение имеет локализация, экстрагируемость, рН-оптимум комплекса ПО и способность синтеза новых компонентов в экстремальных условиях произрастания. Таким образом, ПО-система, по-видимому, участвует в регуляции метаболизма в ходе онтогенеза исследованных растений и играет важную роль в их быстром приспособлении к изменяющимся условиям среды.
Важная роль в обмене веществ в процессах адаптации организма к стресс-факторам отводится КАТ, которая является одним из самых активных ферментов в растениях.
Изменение активности КАТ во многом связано с биологическими особенностями вида и является до некоторой степени показателем реакции растительного организма на комплекс экологических воздействий [16.]. Большинство растений обладают пониженной активностью КАТ на загрязненных участках произрастания, и чем выше устойчивость вида к загрязняющим веществам, тем более высокая стабильность действия этого фермента, и, наоборот, бóльшая степень ингибирования его активности может являться диагностическим признаком слабой устойчивости растений к антропогенным нагрузкам [19.]. Изменения в свойствах фермента могут быть понятны только в контексте общего приспособления всех метаболических реакций клетки к стрессу, т.е. имеют адаптационный характер и дают возможность противостоять неблагоприятным условиям.
Активность ПФО, являющейся одной из терминальных оксидаз растительной клетки, в неблагоприятных и стрессовых условиях среды усиливается, и наряду с ПО участвует в окислении фенольных соединений, которые являются антиоксидантами, и выступают в качестве защитных компонентов растительной клетки [13.]. Поэтому увеличение активности фенолокисляющих ферментов можно рассматривать в качестве одного из индикаторов неустойчивости вида.
Среди неферментативных систем антиоксидантной защиты рассматривается аскорбиновая кислота и каротиноиды, которые способны реагировать с кислородными радикалами и, тем самым, снижать их концентрацию. Загрязненность среды обуславливала снижение ее количества в листьях и корнях. В процессе онтогенеза растений также наблюдалось уменьшение содержания витамина С. Возможно, что различная степень расходования аскорбиновой кислоты в вегетативных органах растений под влиянием загрязняющих веществ обусловлена неодинаковой чувствительностью видов и их онтогенетических групп, разной токсичностью вредных агентов. Поскольку аскорбиновая кислота является ингибитором свободнорадикального окисления, то при действии антропогенных загрязнителей повышается ее расходование на инактивацию свободных радикалов.