Материал: fiza_pischevarenia
1. Пищевой центр. Теории формирования чувства голода, аппетита и насыщения.
Пищевой центр — сложный гипоталамо-лимбико-ретикуло-кортикальный комплекс. Ведущим отделом являются латеральные ядра гипоталамуса. При их поражении наблюдается отказ от пищи, а при электрическом раздражении через вживленные в мозг электроды — повышенное потребление пищи. Эту часть пищевого центра называют центром голода. Разрушение вентромедиальных ядер гипоталамуса приводит к гиперфагии, а их раздражение — к афагии. Считают, что в этих ядрах локализован центр насыщения. Между ним и центром голода установлены реципрокные отношения, т. е. если один центр возбужден, то другой заторможен. Нарушение пищевого поведения происходит также и при поражении лимбической системы, ретикулярной формации и передних отделов коры больших полушарий.
Локальная теория голода: ощущение голода и пищевое поведение вызываются импульсами от периодически сокращающегося свободного от пищи желудка. Эти сокращения желудка назвали «голодными». При наполнении желудка пищей эти сокращения прекращаются и поступающие от желудка импульсы подавляют голод. Акт приема пищи, активация секреции и моторики пищеварительного тракта оказывают тормозное влияние на пищевой центр, вызывая так называемое первичное насыщение. Восстановление метаболизма после всасывания в кровь и депонирования пищевых веществ обеспечивает вторичное насыщение.
Насыщение – это не только снятие чувства голода, но и ощущение удовольствия, полноты в желудке после приема пищи. Постепенно это ощущение угасает. В насыщении существенную роль играют психологические факторы. Глюкостатическая теория: углеводный обмен влияет на аппетит через гипоталамические рецепторы, состояние которых изменяется в зависимости от различия в содержании глюкозы в крови. Аминацидостатическая теория: аминокислоты крови, тормозят пищевой центр. Липостатическая теория: связывает возбуждение пищевого центра с высвобождением липидов из жировых депо. Глюкостатический механизм определяет кратковременные изменения состояния пищевого центра, липостатический — долговременные. Термостатическая теория: избыточное тепло, связанное с метаболическими процессами (и температурой окружающей среды), оказывает тормозное влияние на пищевой центр, является сигналом насыщения. Прием пищи увеличивает теплообразование и участвует в формировании чувства насыщения. Метаболическая теория: гормоны поджелудочной железы (инсулин, глюкагон), гипоталамо-гипофизарной системы (тиролиберин, кортиколиберин, нейротензин, соматотропин), щитовидной железы (тироксин, трийодтиронин, кальцитонин), гормоны пищеварительного тракта (холецистокинин, гастрин), половые гормоны (эстрогены, андрогены).
Аппетит — ощущение, связанное со стремлением человека к определенной пище. Аппетит индивидуально вырабатывается и отражает не столько потребность в пище вообще, сколько потребность в связи со спецификой обмена веществ и дефицитом тех или иных компонентов пищи, индивидуальных и групповых привычек, особенностью национальной кухни и продуктов, которым отдается предпочтение. Избирательный аппетит — это влечение человека к определенному роду пищи, чаще тому, который содержит недостающие организму вещества.
3. Пищеварение в полости рта. Слюноотделение, жевание, глотание, их регуляция.
Поступившая в рот пища раздражает рецепторы полости рта. Тактильные, температурные и болевые рецепторы расположены по всей слизистой оболочке рта, вкусовые — преимущественно во вкусовых почках сосочков языка. Они участвуют в формировании сладкого, кислого, горького и соленого вкуса.
Импульсы от вкусовых рецепторов по афферентным волокнам язычной ветви тройничного, лицевого и языкоглоточного нервов поступают в ЦНС. Эфферентные влияния возбуждают секрецию слюнных, желудочных и поджелудочной желез, желчевыделение, изменяют моторную деятельность пищевода, желудка, проксимального отдела тонкой кишки, влияют на кровоснабжение органов пищеварения, рефлекторно усиливают расходы энергии, необходимой для переработки и усвоения пищи.
Во рту пища в процессе жевания измельчается, смачивается и перемешивается со слюной, растворяется (без чего невозможны оценка вкусовых качеств пищи и ее гидролиз); здесь же формируется ослизненный пищевой комок, предназначенный для глотания. Жевание - процесс механической обработки пищи между верхними и нижними рядами зубов с помощью движения нижней челюсти относительно верхней. Жевательные движения осуществляются сокращениями жевательных и мимических мыщц, мыщц языка.
Импульсы от рецепторов полости рта в основном по волокнам тройничного нерва передаются в сенсорные ядра продолговатого мозга, ядра зрительного бугра, ретикулярной формации, оттуда — в кору большого мозга. В регуляции жевания принимают участие двигательные ядра продолговатого мозга, красное ядро, черное вещество, подкорковые ядра и кора большого мозга. Центр жевания - совокупность управляющих жеванием нейронов различных отделов мозга. Импульсы от него по двигательным волокнам тройничного нерва поступают к жевательным мышцам. При регистрации жевания (мастикациография) выявляются следующие фазы: покоя, введения пищи в рот, ориентировочная, основная, формирования пищевого комка.
Слюна продуцируется тремя парами крупных слюнных желез (околоушная, поднижнечелюстная, подъязычная) и множеством мелких железок языка, слизистой оболочки неба и щек. Из желез по выводным протокам слюна поступает в полость рта.
Слизь слюны выполняет и защитную функцию, покрывая нежную слизистую оболочку рта и пищевода. Слюна содержит муцин и несколько ферментов: α-амилазу, α-глюкозидазу. Гидролиз углеводов, осуществляемый с помощью этих ферментов, из-за кратковременности пребывания пищи в полости рта происходит в основном внутри пищевого комка уже в желудке. Лизоцим слюны обладает высокой бактерицидностью.
Значение слюны в пищеварении состоит в смачивании пищи, что способствует ее измельчению и гомогенизации при жевании; растворении питательных и вкусовых веществ, что важно для раздражения вкусовых рецепторов и действия ферментов слюны; ослизнения принятой и пережеванной пищи, что необходимо для формирования пищевого комка и облегченного его проглатывания.
Прием пищи и связанные с ним факторы рефлекторно возбуждают слюноотделение. Латентный период слюноотделения зависит от силы пищевого раздражителя и возбудимости пищевого центра и составляет 1—30 с. Слюноотделение продолжается весь период еды и почти полностью прекращается вскоре после ее окончания.
Возбуждение от рецепторов полости рта передается в ЦНС по афферентным волокнам тройничного, лицевого, языкоглоточного и блуждающего нервов. Импульсы достигают продолговатого мозга, других отделов мозга, включая кору. Центр слюноотделения расположен в продолговатом мозге и в боковых рогах верхних грудных сегментов спинного мозга. К слюнным железам импульсы следуют по эфферентным парасимпатическим и симпатическим нервным волокнам.
Парасимпатическая иннервация поднижнечелюстной и подъязычной: преганглионарные волокна от верхнего слюноотделительного ядра продолговатого мозга – в составе барабанной струны - до ганглиев этих желез, где переключаются на постганглионарные нейроны – по аксонам достигают гландулоцитов. околоушных желез: преганглионарные волокна от нижнего слюноотделительного ядра продолговатого мозга - в составе языкоглоточного нерва – до ушного ганглия - вторые нейроны – по аксонам в составе ушно-височного нерва - к железам.
Ацетилхолин: большое количество жидкой слюны с высокой концентрацией электролитов и низкой концентрацией муцина. Норадреналин: мало густой слюны, много ферментов и муцина. Симпатическая иннервация слюнных желез осуществляется из боковых рогов II—IV грудных сегментов спинного мозга.
Жевание завершается глотанием — переходом пищевого комка из полости рта в желудок. Глотание возникает в результате раздражения чувствительных нервных окончаний тройничного, гортанных и языкоглоточного нервов. По афферентным волокнам этих нервов импульсы поступают в продолговатый мозг –центр глотания. От него импульсы по эфферентным двигательным волокнам тройничного, языкоглоточного, подъязычного и блуждающего нервов достигают мышц, обеспечивающих глотание.
Рефлекс глотания состоит из трех последовательных фаз: I — ротовой (произвольной); II — глоточной (быстрой, короткой непроизвольной); III — пищеводной (медленной, длительной непроизвольной).
Во время фазы I пищевой комок движениями языка перемещается на его спинку. Произвольными сокращениями языка прижимается к твердому небу и переводится на корень языка за передние дужки.
Во время фазы II раздражение рецепторов корня языка рефлекторно вызывает сокращение мышц, приподнимающих мягкое небо и вызывающих поднятие гортани что препятствует попаданию пищи в дыхательные пути. Движениями языка пищевой комок проталкивается в глотку.
Фазу III глотания составляют прохождение пищи по пищеводу и перевод ее в желудок сокращениями пищевода. Движения пищевода вызываются рефлекторно при каждом глотательном акте. Регуляция моторики пищевода осуществляется в основном эфферентными волокнами блуждающего и симпатического нервов
Вне глотания вход из пищевода в желудок закрыт нижним пищеводным сфинктером. Когда релаксационная волна достигает конечной части пищевода, сфинктер расслабляется и перистальтическая волна проводит через него пищевой комок в желудок. При наполнении желудка тонус кардии повышается, что предотвращает забрасывание содержимого желудка в пищевод.
4. Пищеварение в желудке. Секреторная функция желудка, ее регуляция.
Прием пищи резко увеличивает выделение желудочного сока. Это происходит за счет стимуляции желудочных желез нервными и гуморальными механизмами. Объем и характер секреции, кислотность и содержание в соке пепсинов определяются видом принятой пищи.
Секрецию соляной кислоты обкладочными клетками стимулируют холинергические волокна блуждающих нервов, медиатор которых — ацетилхолин (АХ) — возбуждает М-холинорецепторы мембран гландулоцитов. Путем активации мембранной Na+, К+-АТФазы, увеличения транспорта ионов Са2+ и эффектов повышенного цГМФ, высвобождения гастрина. Эффекты АХ и его аналогов блокируются атропином. Непрямая стимуляция клеток блуждающими нервами опосредуется также гастрином и гистамином.
Гастрин высвобождается из G-клеток, основное количество которых находится в слизистой оболочке пилорической части желудка. Высвобождение гастрина усиливается импульсами блуждающего нерва, а также местным механическим и химическим раздражением этой части желудка. Химическими стимуляторами G-клеток являются продукты переваривания белков. Таким образом, гастрин принимает участие в выделение соляной кислоты в зависимости от величины рН содержимого антрального отдела.
Гистамин, образующийся в ECL-клетках слизистой оболочки желудка стимулирует гландулоциты, влияя на Н2-рецепторы их мембран и и систему аденилатциклаза (АЦ) — цАМФ. Вызывает выделение большого количества сока высокой кислотности, но бедного пепсином. Стимулирующие эффекты гастрина и гистамина зависят от сохранности иннервации желудочных желез блуждающими нервами.
Повышение кислотности содержимого двенадцатиперстной кишки тормозит выделение соляной кислоты железами желудка. Торможение секреции соляной кислоты вызывают секретин, соматостатин, ХЦК, глюкагон, ВИП, тиролиберин, АДГ, кальцитонин, бульбогастрон, кологастрон, окситоцин, серотонин.
Стимуляторами секреции пепсиногена главными клетками являются холинергические волокна блуждающих нервов, гастрин, гистамин, секретин и ХЦК, симпатические волокна, оканчивающиеся на β-адренорецепторах. Усиление секреции пепсиногенов осуществляется механизмами: увеличение переноса ионов Са2+ в клетку и стимуляция Na+, К+-АТФазы; активация системы цГМФ и цАМФ.
Стимуляция секреции слизи мукоцитами осуществляется холинергическими волокнами блуждающих нервов. Гастрин и гистамин умеренно стимулируют мукоциты. Серотонин, соматостатин, адреналин, дофамин — усиливает секрецию слизи.
Три фазы желудочной секреции: Начальная фаза секреции желудка возникает условно-рефлекторно в ответ на раздражение дистантных рецепторов, возбуждаемых видом и запахом пищи, всей обстановкой, связанной с ее приемом. Кроме того, секреция желудка возбуждается безусловно-рефлекторно в ответ на раздражение принимаемой пищей рецепторов полости рта и глотки. Эти рефлексы обеспечивают пусковые влияния на железы желудка. Секреция в мозговую фазу зависит от возбудимости пищевого центра и может легко тормозиться. Так, плохая сервировка стола, неопрятность места приема пищи снижают и тормозят желудочную секрецию. Оптимальные условия приема пищи положительно влияют на желудочную секрецию. Прием в начале еды сильных пищевых раздражителей повышает желудочную секрецию в первую фазу.
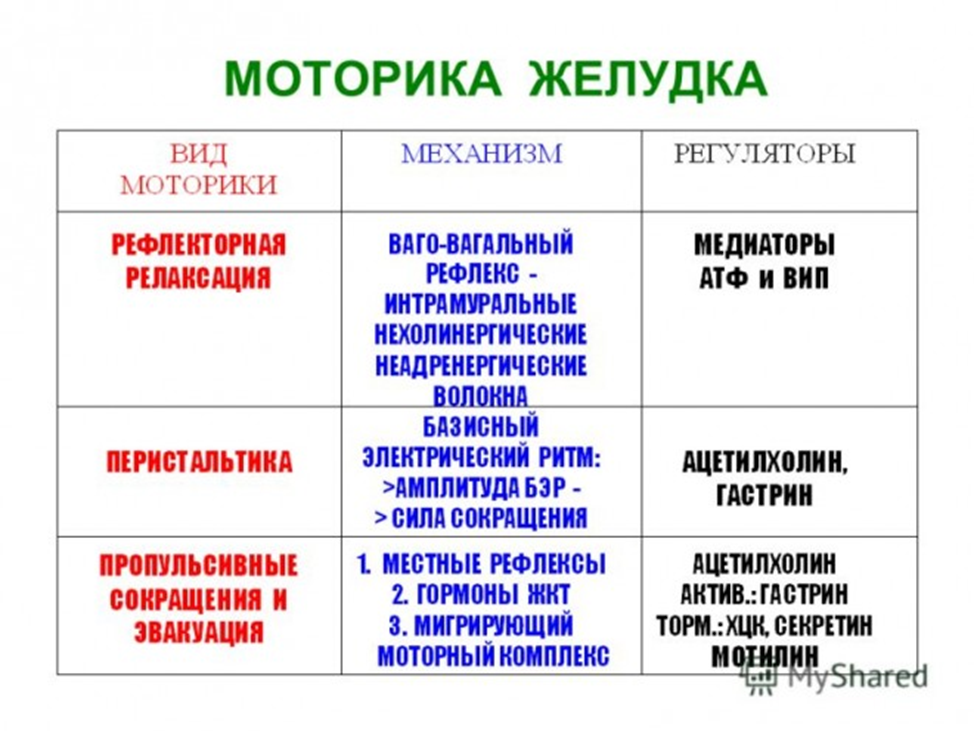
5. Моторная деятельность желудка, ее регуляция. Эвакуация содержимого желудка в 12-перстную кишку.
Во время и в первые минуты после приема пищи наступает пищевая рецептивная релаксация желудка, которая способствует депонированию пищи в желудке и его секреции. Спустя некоторое время в зависимости от вида пищи сокращения желудка начинаются на большой кривизне в непосредственной близости от пищевода, где находится кардиальный водитель ритма. Второй водитель ритма локализован в пилорической части желудка.
Сокращения желудка двух типов: фазовые (тип А) и тонические (тип Б). Волны типа А быстрые, перистальтические, волны типа Б — медленные, тонические. В наполненном пищей желудке возникают три основных вида движений: перистальтические волны, систолические сокращения пилорического отдела тонические, уменьшающие размер полости дна и тела желудка.
В течение первого часа перистальтические волны слабые, в дальнейшем они усиливаются, проталкивая пищу к выходу из желудка. Давление в пилорическом отделе повышается, открывается сфинктер привратника, и порция желудочного содержимого переходит в двенадцатиперстную кишку. Оставшееся (большее) количество его возвращается. Такие движения желудка обеспечивают перемешивание и перетирание пищевого содержимого, его гомогенизацию. Характер, интенсивность, временная динамика моторики зависят от количества и вида пищи, от эффективности ее переваривания в желудке и кишечнике, обеспечивается регуляторными механизмами.
Раздражение блуждающих нервов и выделение АХ усиливают моторику желудка: увеличивают ритм и силу сокращений, ускоряют движение перистальтических волн. Влияния блуждающих нервов могут оказывать и тормозной эффект: рецептивная релаксация желудка, снижение тонуса пилорического сфинктера. Раздражение симпатические нервов и активация α-адренорецепторов тормозят моторику желудка: уменьшают ритм и силу его сокращений, скорость движения перистальтической волны. Стимулирующие α- и β-адренорецепторные влияния (на пилорический сфинктер. Моторику желудка усиливают гастрин, мотилин, серотонин, инсулин. тормозят — секретин, ХЦК, глюкагон, ЖИП, ВИП.
Эвакуация содержимого желудка в двенадцатиперстную кишку Скорость эвакуации пищи из желудка зависит от: объема, состава и консистенции, температуры и рН содержимого желудка, градиента давления между полостями пилорического отдела желудка и двенадцатиперстной кишки, состояния сфинктера привратника, аппетита. Пища, богатая углеводами, быстрее эвакуируется из желудка, чем богатая белками. Жирная пища эвакуируется из него с наименьшей скоростью. Жидкости сразу. Время полной эвакуации смешанной пищи из желудка здорового взрослого человека составляет 6—10 ч.
Регуляция скорости эвакуации содержимого желудка. Осуществляется рефлекторно при активации рецепторов желудка и двенадцатиперстной кишки. Раздражение механорецепторов желудка ускоряет эвакуацию его содержимого, а двенадцатиперстной кишки — замедляет. Из химических агентов, действующих на слизистую оболочку двенадцатиперстной кишки, значительно
Замедляют эвакуацию кислые и гипертонические растворы, 10 % раствор этанола, глюкоза и продукты гидролиза жира, недостаток гидролиза. Желудочная эвакуация «обслуживает» гидролитический процесс в двенадцатиперстной и тонкой кишке и в зависимости от хода его с различной скоростью «загружает» основной «химический реактор» пищеварительного тракта — тонкую кишку.
Регуляторные влияния на моторную функцию гастродуоденального комплекса передаются с интеро- и экстероцепторов через ЦНС и короткие рефлекторные дуги, замыкающиеся в экстра- и интрамуральных ганглиях. В регуляции эвакуаторного процесса принимают участие гастроинтестинальные гормоны, влияющие на моторику желудка и кишечника, изменяющие секрецию главных пищеварительных желез и через нее — параметры эвакуируемого желудочного содержимого и кишечного химуса.